Eritrocito
El eritrocito (del griego ἐρυθρός ‘rojo’, y κύτος ‘bolsa’) —también llamado glóbulo rojo o hematíe—[1] es una célula de la sangre,[2] concretamente la más numerosa. La hemoglobina es uno de sus principales componentes, y su función es transportar el oxígeno hacia los diferentes tipos de tejidos del cuerpo.[3] Los eritrocitos humanos, así como los del resto de mamíferos, carecen de núcleo y de mitocondrias, por lo que deben obtener su energía metabólica a través de la fermentación láctica.[4] La cantidad considerada normal en la especie humana fluctúa entre 4 500 000 (en la mujer) y 5 400 000 (en el hombre) por milímetro cúbico (o microlitro) de sangre,[5] es decir, aproximadamente 1000 veces más que los leucocitos. El exceso de glóbulos rojos se denomina policitemia y su deficiencia se llama anemia.[6][7] Los eritrocitos se utilizan comúnmente en transfusiones en la práctica clínica y se han sugerido como transportadores de fármacos y nanopartículas. El tiempo de vida es de 120 días, posteriormente muere por un proceso llamado apoptosis.[8][9]
| Eritrocito | ||
|---|---|---|
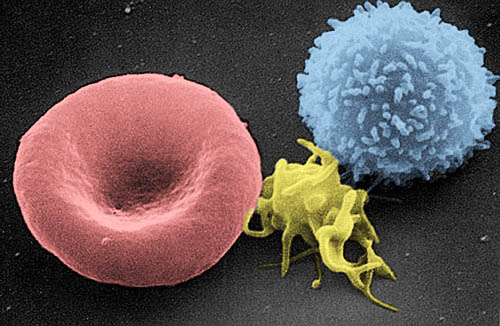 Imagen tomada con un microscopio electrónico en la que se observa, de izquierda a derecha: un glóbulo rojo, una plaqueta y un glóbulo blanco. | ||
| Nombre y clasificación | ||
| Sinónimos |
hematíe. | |
| Latín | Erythrocytus; Haematia | |
| TH | H2.00.04.1.01001 | |
| TH | H2.00.04.1.01001 | |
| Información anatómica | ||
| Sistema | cardiovascular | |
| Producido por | médula ósea | |
|
| ||

Descripción
editarEl glóbulo rojo es un disco bicóncavo de entre 7 y 9 μm de diámetro,[10] de 1 μm de grosor y de 80 a 100 femtolitros de volumen. La célula ha perdido su ARN residual y sus mitocondrias, así como algunas enzimas importantes; por tanto, es incapaz de sintetizar nuevas proteínas o lípidos. Su citoplasma contiene en mayor parte el pigmento hemoglobina, que les concede su característico color rojo (que puede ser más oscuro dependiendo de su oxigenación) y es el responsable del transporte de oxígeno.[11][12]
Ahora bien, esta descripción se aplica a glóbulos rojos de mamíferos, pues en el resto de vertebrados, salvo algunas excepciones, el eritrocito carece de la forma bicóncava y acostumbra ser más grande que el descrito anteriormente. Esto se debe a que los glóbulos rojos del resto de vertebrados todavía poseen núcleo.
Los eritrocitos derivan de las células madre comprometidas denominadas hemocitoblasto. La eritropoyetina, una hormona de crecimiento producida en los tejidos renales, estimula la eritropoyesis (es decir, la formación de eritrocitos) y es responsable de mantener una masa eritrocitaria en un estado constante. Los eritrocitos, al igual que los leucocitos, tienen su origen en la médula ósea.
La concentración eritrocitaria varía según el sexo, la edad y la ubicación geográfica. Se encuentran concentraciones más altas de eritrocitos en zonas de gran altitud, en varones y en recién nacidos. Las disminuciones por debajo del rango de referencia generan un estado patológico denominado anemia. Esta alteración provoca hipoxia tisular. El aumento de la concentración de eritrocitos (policitemia) es menos común.
La hemólisis es la destrucción de los hematíes envejecidos y sucede en los macrófagos del bazo e hígado. Los elementos esenciales, globina y hierro, se conservan y vuelven a usarse. La fracción hemo de la molécula se cataboliza a bilirrubina y a biliverdina, y finalmente se excreta a través del tracto intestinal. La rotura del eritrocito a nivel intravascular libera hemoglobina directamente a la sangre, donde la molécula se disocia en dímeros α y β, los cuales se unen a la proteína de transporte, haptoglobina. Esta transporta los dímeros al hígado, donde posteriormente son catabolizados a bilirrubina y se excretan.
En mamíferos
editarLos glóbulos rojos de muchos mamíferos no poseen núcleo cuando llegan a la madurez, es decir, pierden su núcleo celular y por lo tanto su ADN; los anfibios, reptiles y aves tienen hematíes con núcleo. Los eritrocitos también pierden sus mitocondrias y utilizan la glucosa para producir energía mediante el proceso de glucólisis seguido por la fermentación láctica.
Los glóbulos rojos son producidos continuamente en la médula ósea de los huesos largos, aunque en el embrión, el hígado es el principal productor de eritrocitos. El bazo actúa como reservorio de eritrocitos, pero su función es algo limitada en el ser humano. Sin embargo, en otros mamíferos, como los perros y los caballos, el bazo libera grandes cantidades de eritrocitos en momentos de estrés. Algunos atletas han tratado de explotar esta función del bazo tratando de liberar sus reservas de hematíes mediante fármacos, pero esta práctica pone en riesgo al aparato cardiovascular, dado que este no está preparado para soportar sangre cuya viscosidad sea superior a la considerada normal.
En seres humanos
editarLos eritrocitos tienen una forma oval, bicóncava, aplanada, con una depresión en el centro. Este diseño es el óptimo para el intercambio de oxígeno con el medio que lo rodea, pues les otorga flexibilidad para poder atravesar los capilares, donde liberan la carga de oxígeno. El diámetro de un hematíe típico es de 6-8 µm. Los glóbulos rojos contienen hemoglobina, que se encarga del transporte de oxígeno y del dióxido de carbono. Asimismo, es el pigmento que le da el color rojo a la sangre.
Valores considerados normales en adultos
editar- Mujeres: 4,7 ± 0,7 x 106/μL (microlitro) de sangre
- Hombres: 5,3 ± 0,8 x 106/μL (microlitro) de sangre
Maduración
editarDada la necesidad constante de reponer los glóbulos rojos, las células eritropoyéticas de la médula ósea se cuentan entre las de crecimiento y reproducción más rápidas de todo el cuerpo. Por tanto, como cabría esperar, su maduración y producción resultan muy afectadas en casos de deficiencias nutricionales importantes.
Para la maduración final del hematíe se necesitan en particular dos vitaminas: la vitamina B12 y el ácido fólico. Ambas son esenciales para la síntesis del ADN porque las dos, de forma diferente, resultan necesarias para la formación de trifosfato de timidina, uno de los componentes esenciales del ADN. Por lo tanto, la carencia de vitamina B12 o de ácido fólico originan una disminución de la producción de ADN y, en consecuencia, determina un fracaso de la maduración y división nuclear.
Asimismo, las células eritroblásticas de la médula ósea, además de no proliferar con rapidez, originan sobre todo eritrocitos de mayor tamaño que el normal denominados macrocitos, con una membrana muy delgada, irregular y oval, en lugar del disco bicóncavo habitual. Estas células mal formadas, tras entrar en la sangre circulante, transportan oxígeno con normalidad, pero debido a su fragilidad, su vida se acorta de la mitad a una tercera parte. Por eso, se dice que el déficit de vitamina B12 o de ácido fólico produce un fracaso de la maduración eritropoyética.
Existen otras causas que alteran la maduración de los glóbulos rojos, como la deficiencia de hierro y otras anomalías genéticas que conducen a la producción de hemoglobinas anormales. Todos estos problemas conducirán a alteraciones de los eritrocitos, por alteración de la membrana, el citoesqueleto u otros.
Etapas de desarrollo morfológico
editarLas etapas de desarrollo morfológico de la célula eritroide incluyen (en orden de madurez creciente) las siguientes etapas:
- Célula madre pluripotencial.
- Célula madre multipotencial.
- Célula progenitora o CFU-S (unidad formadora de colonias del bazo).
- BFU-E (unidad formadora de brotes de eritrocitos).
- CFU-E (unidad formadora de colonias de eritrocitos), que luego formará los proeritroblastos.
- Proeritroblasto: Célula grande de citoplasma abundante, núcleo grande con cromatina gruesa, nucléolos no muy bien definidos (20-25 micras).
- Eritroblasto basófilo: Más pequeño que el anterior (16-18 micras), citoplasma basófilo, cromatina gruesa y grumosa, aquí se inicia la formación de la hemoglobina.
- Eritroblasto policromatófilo: Mide 10-12 micras, el citoplasma empieza a adquirir un color rosa por la presencia de hemoglobina, aquí se presenta la última fase mitótica para la formación de hematíes, no posee nucléolos y la relación núcleo/citoplasma es de 4:1.
- Eritroblasto ortocromático: Mide 8-10 micras, tiene cromatina compacta y el núcleo empieza a desaparecer.
- Reticulocito: Casi diferenciado en eritrocitos maduros. La presencia de reticulocitos en sangre periférica) indica el buen funcionamiento de la médula ósea.
- Eritrocito, finalmente, cuando ya carece de núcleo y mitocondrias. Tiene capacidad de transporte (gases, hormonas, medicamento, etc.).
A medida que la célula madura, la producción de hemoglobina aumenta, lo que genera un cambio en el color del citoplasma en las muestras de sangre teñidas con la tinción de Wright, de azul oscuro a gris rojo y rosáceo. El núcleo paulatinamente se vuelve picnótico, y es expulsado fuera de la célula en la etapa ortocromática.
La membrana del eritrocito en un complejo bilipídico–proteínico, el cual es importante para mantener la deformabilidad celular y la permeabilidad selectiva. Al envejecer la célula, la membrana se hace rígida, permeable y el eritrocito es destruido en el bazo. La vida media promedio del hematíe normal es de 100 a 120 días.[8]
Composición de la membrana
editarLa membrana del eritrocito tiene varias funciones que ayudan en la regulación superficial de la deformación, flexibilidad, adhesión a otras células y reconocimiento inmunológico. Estos atributos son altamente dependientes de su composición, lo cual define sus propiedades. La membrana del eritrocito está compuesta de tres capas: el glicocálix al exterior, que es rico en carbohidratos; la bicapa lipídica que contiene varias proteínas transmembranales además de sus constituyentes lipídicos principales; y el citoesqueleto membranal, una red estructural de proteínas localizado en la superficie interna de la bicapa lipídica. La mitad de la masa de la membrana del eritrocito en humanos y la mayoría de los mamíferos son proteínas, la otra mitad son lípidos, principalmente fosfolípidos y colesterol.
Lípidos de membrana
editarLa membrana del eritrocito está compuesta por una bicapa lipídica, similar a la que se encuentra prácticamente en todas las células humanas. Esta bicapa lipídica está compuesta de colesterol y fosfolípidos en proporciones iguales en peso. La composición lipídica es importante debido a que define muchas propiedades físicas como la permeabilidad y la fluidez. Además, la actividad de varias proteínas de membrana es regulada por la interacción con los lípidos de la bicapa. A diferencia del colesterol que se encuentra distribuido de manera uniforme entre las monocapas interna y externa, los 5 fosfolípidos principales están dispuestos de forma asimétrica:
En la monocapa externa
En la monocapa interna
La distribución asimétrica de los fosfolípidos en la bicapa es el resultado de la función de algunas proteínas transportadoras de fosfolípidos tanto dependientes como independientes de energía. Las flipasas son proteínas que mueven fosfolípidos de la monocapa externa a la interna, mientras que las llamadas flopasas hacen la operación inversa, en contra del gradiente de concentración de manera dependiente de energía. Además, están las proteínas escramblasas que mueven fosfolípidos en ambas direcciones al mismo tiempo, por sus gradientes de concentración e independientes de energía. Todavía está en discusión la identidad de las proteínas de mantenimiento de membrana en los hematíes.
El mantenimiento de la distribución asimétrica de fosfolípidos en la bicapa es crítica para la integridad y funcionalidad de la célula debido a varias razones:
- Los macrófagos reconocen y fagocitan eritrocitos que tienen expuesto la fosfatidilserina en la superficie externa. Por lo cual mantener a la fosfatidilserina en la monocapa interna es esencial para la supervivencia de la célula en sus encuentros frecuentes con macrófagos del sistema retículo-endotelial, especialmente en el bazo.
- Destrucción prematura de eritrocitos talasémicos y falsiformes han sido ligados a la desorganización de la asimetría lipídica llevando a la exposición de la fosfatidilserina en la monocapa externa.
- Una exposición de fosfatidilserina puede potenciar la adhesión de los glóbulos rojos a las células endoteliales vasculares, evitando eficazmente el tránsito normal por la microvasculatura. Por ello es importante mantener a la fosfatidilserina en la monocapa interna de la bicapa para asegurar un flujo sanguíneo normal en micro-circulación.
- La fosfatidilserina y el fosfatidilinositol-4,5-bifosfato (PtdIns(4,5)P2) pueden regular la función mecánica de la membrana, debido a sus interacciones con las proteínas del citoesqueleto como la espectrina y la proteína 4.1R. Estudios recientes señalan que la unión de la espectrina a fosfatidilserina promueve la estabilidad mecánica en la membrana. El PtdIns(4,5)P2 mejora la unión de la proteína 4.1R a la glicoforina C pero disminuye su interacción con la banda de proteína 3, y de este modo puede modular la unión de la bicapa al citoesqueleto.
La presencia de estructuras especializadas llamadas balsas lipídicas en la membrana de los eritrocitos han sido descritas en estudios recientes. Estas estructuras ricas en colesterol y esfingolípidos están asociados a proteínas de membrana específicas, como la proteína G.[13][14]
Metabolismo energético
editarEl metabolismo del glóbulo rojo es limitado, debido a la ausencia de núcleo, mitocondria y otros orgánulos subcelulares. Aunque la unión, el transporte y la liberación de oxígeno y dióxido de carbono es un proceso pasivo que no requiere energía, existe una variedad de procesos metabólicos dependientes de energía que son esenciales para la viabilidad de la célula.
Las vías metabólicas más importantes para el eritrocito maduro necesitan glucosa como sustrato. Estas vías se refieren a:
- glucólisis
- ruta de la pentosa fosfato
- vía de la hemoglobina reductasa
- ciclo de Rapoport–Luebering
Estas vías contribuyen con energía, al mantener:
- el potasio intracelular alto, el sodio intracelular bajo y un calcio intracelular muy bajo (bomba de cationes);
- hemoglobina en forma reducida;
- elevados niveles de glutatión reducido;
- integridad y deformabilidad de la membrana.
Vía Embden–Meyerhof o glucólisis anaeróbica
editarProporciona ATP para la regulación de la concentración intracelular de cationes (Na+, K+, Ca2+, Mg2+) a través de bombas de cationes. El eritrocito obtiene energía en forma de ATP del desdoblamiento de la glucosa por esta vía. Los eritrocitos normales no tienen depósitos de glucógeno, dependen por completo de la glucosa ambiental para la glucólisis. La glucosa penetra a la célula mediante difusión facilitada, un proceso que no consume energía. Es metabolizada a lactato, donde produce una ganancia neta de dos moles de ATP por un mol de glucosa.
Ciclo de las pentosas
editarProporciona nicotinamida-adenina dinucleótido fosfato y glutatión reducido para reducir oxidantes celulares. Aproximadamente el 5 % de la glucosa celular ingresa a la vía oxidativa de las pentosas, un sistema auxiliar para producir coenzimas reducidas. El glutatión reducido protege a la célula contra muchas lesiones producidas por agentes oxidantes permanentes. Los oxidantes dentro de la célula oxidan los grupos sulfhidrilo (-SH) de la hemoglobina, a menos que los oxidantes sean reducidos por el glutatión reducido. Es por esto que es crucial en el eritrocito la función de esta vía.
Vía de la hemoglobina reductasa
editarProtege a la hemoglobina de la oxidación vía la NADH y metahemoglobina reductasa. Se trata de una vía alterna a la vía Embden–Meyerhof, esencial para mantener al hierro hemo en el estado reducido Fe++. La hemoglobina con el hierro en estado férrico, Fe3+, es conocida como metahemoglobina. Esta forma de hemoglobina no logra combinarse con el oxígeno. La metahemoglobina reductasa, en unión con el NADH producido por la vía Embden–Meyerhof, protege al hierro hemo de la oxidación. Sin este sistema, el 2 % de la metahemoglobina formada todos los días se elevaría, con el tiempo, a un 20-40 %, con lo que se limitaría gravemente la capacidad transportadora de oxígeno en la sangre. Los medicamentos oxidantes pueden interferir con la metahemoglobina reductasa y producir valores aún más elevados de metahemoglobina. Esto provoca cianosis.
Ciclo de Rapoport–Luebering
editarEste ciclo es parte de la vía Embden–Meyerhof, y tiene por finalidad evitar la formación de 3–fosfoglicerato y ATP. El BPG (2,3-bisfosfoglicerato) está presente en el glóbulo rojo en una concentración de un mol BPG/mol de hemoglobina, y se une con fuerza a la desoxihemoglobina, con lo que la hemoglobina se mantiene en estado desoxigenado y se facilita la liberación de oxígeno. El incremento en la concentración de difosfoglicerato facilita la liberación de oxígeno a los tejidos mediante la disminución en la afinidad de la hemoglobina por el oxígeno. De esta manera, el hematíe cuenta con un mecanismo interno para la regulación del aporte de oxígeno a los tejidos.
Hemoglobina
editarEs un pigmento especial que da a los eritrocitos su color rojo característico. Su molécula posee hierro, y su función es el transporte de oxígeno. Está presente en todos los animales, excepto en algunos grupos de animales inferiores. Participa en el proceso por el que la sangre lleva los nutrientes necesarios hasta las células del organismo y conduce sus productos de desecho hasta los órganos excretores. También transporta el oxígeno desde los pulmones (o desde las branquias, en los peces), donde la sangre lo capta, hasta los tejidos del cuerpo.
Cuando la hemoglobina se une al oxígeno para ser transportada hacia los órganos del cuerpo, se llama oxihemoglobina. Cuando la hemoglobina se une al CO2 para ser eliminada por la espiración, que ocurre en los pulmones, recibe el nombre de Carboaminohemoglobina (también se denomina desoxihemoglobina a la hemoglobina cuándo no está unida al oxígeno). Si la hemoglobina se une al monóxido de carbono (CO), se forma entonces un compuesto muy estable llamado carboxihemoglobina, que tiene un enlace muy fuerte con el grupo hemo de la hemoglobina e impide la captación del oxígeno, con lo que se genera fácilmente una anoxia que conduce a la muerte.
La hemoglobina también transporta productos residuales y el dióxido de carbono de vuelta a los tejidos. Menos del 2 % total del oxígeno, y la mayor parte del CO2, son mantenidos en solución en el plasma sanguíneo. La hemoglobina representa el 35 % del peso del eritrocito. Un compuesto relacionado, la mioglobina, actúa como almacén de oxígeno en las células musculares.
Véase también
editarReferencias
editar- ↑ Real Academia Española. «hematíe». Diccionario de la lengua española (23.ª edición).
- ↑ Real Academia Española. «glóbulo rojo». Diccionario de la lengua española (23.ª edición).
- ↑ X, Fuentes Arderiu (1998). Bioquímica clínica y patología molecular. II. Reverte. ISBN 978-84-291-1855-1. Consultado el 27 de noviembre de 2019.
- ↑ Campbell, Neil A.; Reece, Jane B. (2007). Biología. Ed. Médica Panamericana. ISBN 978-84-7903-998-1. Consultado el 27 de noviembre de 2019.
- ↑ Donnersberger, Anne B.; Lesak, Anne E. (2002-12). Libro de Laboratorio de Anatomía y Fisiología. Editorial Paidotribo. ISBN 978-84-8019-652-9. Consultado el 27 de noviembre de 2019.
- ↑ Arturo M. Musso: Eritrocitos y eritrocitopatías. Archivado el 20 de septiembre de 2018 en Wayback Machine. Hematología, Vol.18 N° 2: 151-155, mayo - agosto 2014.
- ↑ Guyton, A. C. Tratado de fisiología médica. ISBN 0-7216-4394-9
- ↑ a b «eritrocito». Real Academia Nacional de Medicina de España.
- ↑ Brenner, Jacob S.; Pan, Daniel C.; Myerson, Jacob W.; Marcos-Contreras, Oscar A.; Villa, Carlos H.; Patel, Priyal; Hekierski, Hugh; Chatterjee, Shampa et al. (11 de julio de 2018). «Red blood cell-hitchhiking boosts delivery of nanocarriers to chosen organs by orders of magnitude». Nature Communications (en inglés) 9 (1). ISSN 2041-1723. doi:10.1038/s41467-018-05079-7. Consultado el 12 de julio de 2018.
- ↑ Welsch, Ulrich; Sobotta, Johannes (2008). Histología. Ed. Médica Panamericana. ISBN 978-84-9835-178-1. Consultado el 27 de noviembre de 2019.
- ↑ Mary Louise Turgeon (2004). Clinical Hematology: Theory and Procedures. Lippincott Williams & Wilkins. p. 100. ISBN 9780781750073.
- ↑ McLaren CE, Brittenham GM, Hasselblad V (abril de 1987). «Statistical and graphical evaluation of erythrocyte volume distributions». Am. J. Physiol. 252 (4 Pt 2): H857-66. PMID 3565597.
- ↑ Mohandas N, Gallagher PG (noviembre de 2008). «Red cell membrane: past, present, and future». Blood 112 (10): 3939-48. PMC 2582001. PMID 18988878. doi:10.1182/blood-2008-07-161166.
- ↑ Rodi PM, Trucco VM, Gennaro AM (junio de 2008). «Factors determining detergent resistance of erythrocyte membranes». Biophysical Chemistry 135 (1–3): 14-8. PMID 18394774. doi:10.1016/j.bpc.2008.02.015.
Enlaces externos
editarWikcionario tiene definiciones y otra información sobre eritrocito.
Wikimedia Commons alberga una categoría multimedia sobre Eritrocito.
Datos: Q37187