Músculo esquelético
| Tecido muscular estriado esquelético | |
|---|---|
 | |
| Esquema do músculo esquelético | |
| Latín | textus muscularis striatus skeletalis (TH 2.00.05.2.00002) |
O músculo esquelético ou tecido muscular estriado esquelético é un tipo de músculo estriado que está baixo o control do sistema nervioso somático, polo que se controla voluntariamente. É un dos tres grandes tipos de tecido muscular; os outros dous son o músculo cardíaco (tamén de tipo estriado) e o músculo liso (non estriado). Como indica o seu nome, a maioría dos músculos esqueléticos están unidos aos ósos por feixes fibrosos de coláxeno chamados tendóns. A súa misión é realizar os movementos voluntarios do corpo.
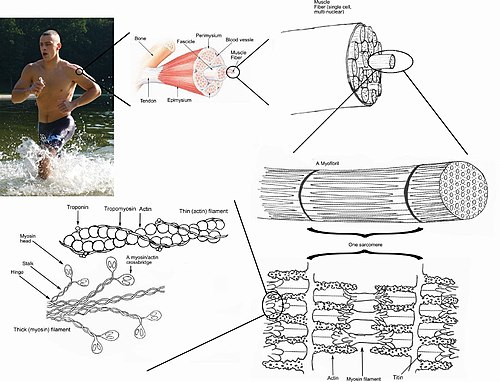
O músculo esquelético está feito de compoñentes individuais denominados miocitos, "células musculares", "fibras musculares" ou miofibras. Os miocitos son células longas cilíndricas e multinucleadas, que se formaron pola fusión de mioblastos durante o desenvolvemento (un tipo de células proxenitoras embrionais) durante un proceso chamado mioxénese.
As miofibras están á súa vez cheas de miofibrilas. As miofibrilas están compostas por filamentos proteicos de actina e miosina repetidos en unidades chamadas sarcómeros, a unida funcional básica da fibra muscular. O sarcómero é responsable da estriación da fibra muscular esquelética (debido á superposición ordenada de filamentos en bandas) e constitúe a maquinaria básica necesaria para a contracción muscular. Os músculos son órganos formados por moitos feixes de fibras musculares unidas por tecido conxuntivo.
Fibras musculares esqueléticas
[editar | editar a fonte]Estrutura das fibras musculares
[editar | editar a fonte]
- Artigo principal: Miocito.
As fibras musculares individuais fórmanse durante o desenvolvemento a partir da fusión de varias células inmaturas indiferenciadas coñecidas como mioblastos orixinando células multinucleadas longas e cilíndricas. A diferenciación neste estado complétase principalmente antes do nacemento e despois as células continúan crecendo en tamaño. O músculo esquelético mostra un patrón de bandas distintivo visto ao microscopio debido á disposición ordenada dos compoñentes do seu citoesqueleto no citoplasma das fibras musculares. As principais proteínas citoplásmicas son miosina e actina, tamén chamadas filamentos "grosos" e "finos", respectivamente, os cales forman unidades repetidas chamadas sarcómeros. A interacción da miosina e a actina é responsable da contracción muscular.
Cada orgánulo e macromolécula dunha fibra muscular está colocado de maneira que permita a súa función. A membrana plasmática denomínase sarcolema e o citoplasma coñécese como sarcoplasma. No sarcoplasma hai conxuntos de miofibrilas. As miofibrilas son longos feixes de proteínas de aproximadamente 1 micron de diámetro que conteñen miofilamentos. Os mionúcleos son aplanados e periféricos, xa que están presionados contra a cara interna do sarcolema. Estre as miofibrilas están as mitocondrias.
Aínda que as fibras musculares non teñen un retículo endoplasmático liso normal, presentan un tipo especial moi característico chamado retículo sarcoplásmico. O retículo sarcoplásmico rodea as miofibrilas e mantén unha reserva de ións calcio que cómpre para causar unha contracción muscular. Cun espazado periódico, presenta sacos con fondo dilatado coñecidos como cisternas terminais. Estas cruzan a fibra muscular dun lado a outro. Entre dúas cisternas terminais hai uns repregamentos tubulares invaxinados do sarcolema chamados túbulos transversos ou túbulos T. Os túbulos T son as vías a través das cales os potenciais de acción indican ao retículo sarcoplásmico que ten que liberar o calcio, causando a contracción muscular. En conxunto, dúas cisternas terminais e un túbulo transverso forman unha tríade.[1]
Outro grupo de células, as células miosatélites encóntrase entre a lámina basal e o sarcolema das fibras musculars. Estas células son normalmente quiescentes pero poden ser activadas polo exercicio ou unha patoloxía e proporcionan mionúcleos adicionais para o crecemento muscular e a reparación.
Tipos de fibras
[editar | editar a fonte]
Empréganse moitos métodos para a tipificación das fibras musculares, e é corrente que haxa unha certa confusión entre os métodos entre os non expertos. Dous métodos que se confunden habitualmente son a marcaxe histoquímica para a actividade ATPase da miosina e a marcaxe inmunohistoquímica para o tipo de miosina de cadea pesada (MHC). A actividade ATPase da miosina denomínase normalmente e de forma correcta simplemente como "tipo de fibra", e orixínase dos ensaios directos da actividade ATPase en varias condicións (por exemplo, de pH).[2] A marcaxe da cadea pesada da miosina denomínase con máis exactitude "tipo de fibra MHC", por exemplo "fibras MHC IIa", e orixínase da determinación de diferentes isoformas de MHC.[2] Estes métodos están moi relacionados fisioloxicamente, xa que o tipo MHC é o principal determinante da actividade ATPase. Póren, ningún destes métodos de tipificación é de natureza directamente metabólica, e non salienta directamente a capacidade oxidativa ou glicolítica da fibra. Cando se fai referencia xenericamente a fibras de "tipo I" ou de "tipo II", isto máis exactamente fai referencia á suma de tipos de fibras numéricos (I fronte a II) avaliadas pola marcaxe da actividade ATPase da miosina (por exemplo, fibras de "tipo II" quere dicir de tipo IIA + tipo IIAX + tipo IIXA... etc.).
A táboa que segue mostra a relación que hai entre estes dous métodos, limitada aos tipos de fibras humanas. Nótese a escritura con maiúscula de subtipos utilizada na tipificación de fibras pola ATPase fronte á tipificación MHC en minúscula, e que algúns tipos clasificados pola actividade ATPase conteñen en realidade moitos tipos MHC. Ademais, en humanos non se expresan os subtipos B nin b por ningún dos métodos.[3] Os primeiros investigadores crían que os humanos expresaban un tipo MHC IIb, o que levou a que na clasificación pola ATPase se incluíse IIB. Porén, investigacións posteriores indicaron que a MHC IIb humana era de feito o tipo IIx,[3] polo que é mellor denominar a IIB como IIX. O tipo IIb exprésase noutros mamíferos, polo que se pode atopar (correctamente) na literatura (xunto con IIB). Os tipos de fibras non humanas inclúen as verdadeiras fibras IIb, IIc, IId etc.
| Tipos pola ATPase | Cadeas pesadas MHC |
|---|---|
| Tipo I | MHC Iβ |
| Tipo IC | MHC Iβ > MHC IIa |
| Tipo IIC | MHC IIa > MHC Iβ |
| Tipo IIA | MHC IIa |
| Tipo IIAX | MHC IIa > MHC IIx |
| Tipo IIXA | MHC IIx > MHC IIa |
| Tipo IIx | MHC IIx |
Outros tipos de métodos de tipificación das fibras están delineados menos formalmente, e existen en máis dun espectro. Tenden a estar enfocados máis ás capacidades metabólicas e funcionais (é dicir, oxidativa fronte a glicolítica, de tempo de contracción rápido fronte a lento). Como xa se indicou, a tipificación de fibras pola ATPase ou a MHC non mide directamente ou determina esas características. Porén, moitos dos métodos están ligados mecanicamente, mentres que outros están correacionados in vivo.[6][7] Por exemplo, os tipos de fibras pola ATPase están relacionados coa velocidade de contracción, porque unha alta actividade de ATPase permite uns ciclos de formación de pontes cruzadas máis rápidos.[2] Aínda que a actividade de ATPase é só un compoñente da velocidade de contracción, as fibras de tipo I son "lentas", en parte, porque teñen velocidades baixas de actividade de ATPase en comparación coa fibras de tipo II. Con todo, medir a velocidade de contracción non é o mesmo que un tipificado das fibras pola ATPase.
Debido a estes tipos de relacións, as fibras de tipo I e II teñen propiedades metabólicas, contráctiles e de unidade motora relativamente diferentes. A táboa que segue diferencia estes tipos de propiedades. Porén, debe terse presente que este tipo de propiedades (aínda que dependan en parte das propiedades das fibras individuais) tenden a ser só relevantes e a ser medidas a nivel de unidade motora e non de fibra individual.[2]
| Propiedades | Fibras de tipo I | Fibras de tipo IIA | Fibras de tipo IIX |
|---|---|---|---|
| Tipo de unidade motora | Oxidativa lenta (SO) | Oxidativa/Glicolítica rápida (FOG) | Glicolítica rápida (FG) |
| Velocidade de contracción | Lenta | Rápida | Rápida |
| Forza de contacción | Pequena | Media | Grande |
| Resistencia á fatiga | Alta | Alta | Baixa |
| Contido de glicóxeno | Baixo | Alto | Alto |
| Irrigación de capilares | Abondosa | Abondosa | Escasa |
| Mioglobina | Alta | Alta | Baixa |
| Cor vermella | Escura | Escura | Clara |
| Densidade de mitocondrias | Alta | Alta | Baixa |
| Densidade de capilares | Alta | Intermedia | Baixa |
| Capacidade encimática oxidativa | Alta | Intermedia-alta | Baixa |
| Amplitude da liña Z | Intermedia | Ampla | Estreita |
| Actividade de ATPase alcalina | Baixa | Alta | Alta |
| Actividade de ATPase ácida | Alta | Media-alta | Baixa |
- Cor das fibras
Tradicionalmente, as fibras foron clasificadas segundo a súa cor, a cal reflectía o seu contido en mioglobina. As fibras de tipo I eran vermellas debido aos seus altos niveis de mioglobina. As fibras musculares vermellas adoitan ter máis mitocondrias e unha maior densidade local de capilares. Estas fibras teñen máis resistencia e tardan máis en fatigarse porque usan un metabolismo oxidativo para xerar ATP. As fibras de tipo II son menos oxidativas e son brancas debido ao seu nivel relativamente baixo de mioglobina e a súa dependencia dos encimas glicolíticos.
- Velocidade de contracción
As fibras poden tamén clasificarse pola súa capacidade de contracción, en fibras de contracción rápida e lenta. Esta característica solápase en grande medida, pero non totalmente, coas clasificacións baseadas na cor, actividade ATPase, ou MHC.
Algúns autores definen a fibra de contracción rápida como aquela na que a miosina pode hidrolizar moi rapidamente o ATP. Isto inclúe na definición principalmente as fibras pola clasificación da ATPase de tipo II e as MHC de tipo II. Porén, as fibras de contracción rápida tamén presentan unha maior capacidade de transmisión electroquímica dos potenciais de acción e unha rápida liberación e captación de calcio no retículo sarcoplásmico. As fibras de contracción rápida dependen dun sistema glicolítico de curto prazo ben desenvolvido para a transferencia de enerxía e poden contraerse e xerar tensión a unha velocidade dúas ou tres veces maior que as fibras de contracción rápida. Os músculos de contracción rápida son moito mellores á hora de xerar curtas explosións de forza ou velocidade que os músculos lentos, pero fatíganse máis rapidamente.[8]
As fibras de contracción lenta xeran enerxía para a resíntese do ATP por medio dun sistema de longo prazo de transferencia de enerxía aerobio. Entre elas están as fibras da clasificación pola ATPase de tipo I e as MHC de tipo I. Adoitan ter un baixo nivel de actividade de ATPase, unha velocidade de contracción máis lenta e unha capacidade glicolítica menos desenvolvida. Conteñen grandes volumes de mitocondrias, e altos niveis de mioglobina que lles dan cor vermella. Teñen unha alta concentración de encimas mitocondriais, polo que son máis resistentes á fatiga. Os músculos de contracción lenta dispáranse máis lentamente que os de contracción rápida, pero poden contraerse durante un período de tempo maior antes de fatigarse.[8]
- Distribución dos distintos tipos
Os músculos do corpo considerados individualmente adoitan estar formados por unha mestura de varios tipos de fibras, pero as súas proporcións varían dependendo das accións que ten que realizar dito músculo e da especie. Por exemplo, nos humanos, o cuadríceps contén ~52% de fibras de tipo I, mentres que o sóleo ten ~80% de tipo I.[9] O músculo orbicular dos ollos ten só o ~15% de fibras de tipo I.[9] A diferenza do anterior, as unidades motoras dun músculo presentan unha mínima variación entre as fibras que compoñen cada unidade. Este feito é o que fai que o tamaño sexa o principal para a viabilidade dunha unidade motora de recrutamento.
Pensábase tradicionalmente que o número total de fibras musculares esqueléticas non cambiaba. Porén, crese que non hai diferenzas en sexo ou idade na distribución das fibras, e os tipos relativos de fibras varían considerablemente dun músculo a outro e dunha persoa a outra. Os homes ou mulleres sedentarios (e os nenos pequenos) teñen un 45% de fibras de tipo II e un 55% de tipo I. Os deportistas de alto nivel adoitan ter patróns de distribución de fibras determinados; por exemplo, os atletas de probas de resistencia mostran un maior nivel de fibras de tipo I. Os atletas de probas de velocidade explosiva (sprints), necesitan grandes cantidades de fibras de tipo IIb (da clasificación tradicional das fibras). Os atletas de probas de media distancia teñen aproximadamente unha distribución igual de ambos os tipos de fibras. Este tamén adoita ser o caso dos atletas de probas de potencia muscular, como os lanzadores e saltadores. Propúxose que certos tipos de exercicios podían inducir cambios nos tipos de fibras esqueléticas presentes nos músculos.[10] Crese que se unha persoa realiza exercicios de resistencia durante un período prolongado da súa vida, algunhas das fibras de tipo IIb transfórmanse en fibras de tipo IIa, aínda que non hai consenso sobre este aspecto. Pode ocorrer tamén que as fibras de tipo IIb experimenten unha potenciación da súa capacidade oxidativa despois dun adestramento prolongado en resistencia de alta intensidade, o cal vai levalas ata un nivel no que poidan realizar o metabolismo oxidativo con igual efectividade ca as fibras de contracción lenta das persoas non adestradas. Isto produciríase por un incremento no tamaño das mitocondrias e no número e os cambios asociados relacionados e non por un cambio de tipo de fibras.
Arquitectura e organización das fibras
[editar | editar a fonte]A arquitectura do músculo refírese á disposición das fibras musculares en relación co eixe de xeración de forza do músculo. Este eixe é unha liña hipotética desde a orixe do músculo ata a súa inserción. Para algúns músculos lonxitudinais, como o bíceps braquial, este é un concepto relativamente simple. Para outros, como o recto femoral ou o deltoide, é máis complicado. Aínda que as fibras musculares dun fascículo son paralelas unhas a outras, os fascículos poden variar na súa relación con outros fascículos ou cos seus tendóns.[11] As diferentes disposicións das fibras orixinan tres grandes clases de arquitectura no músculo esquelético: lonxitudinal, pennada (ou unipennada), e multipennada.[12] Debido ás diferentes arquitecturas que poden ter, a tensión que pode crear un músculo entre os seus tendóns varía máis do que se esperaría simplemente polo seu tamaño e tipo de fibras que o forman.
- Arquitectura lonxitudinal
Os fascículos dos músculos fusiformes, paralelos ou dispostos lonxitudinalmente, corren paralelamente ao eixe de xeración da forza, polo que en conxunto estes músculos funcionan de xeito similar ás fibras musculares grandes sinxelas.[11] Hai variacións, e utilízanse a miúdo diferentes termos máis especificamente. Por exemplo, fusiforme indica unha arquitectura lonxitudinal cunha barriga muscular máis gorda (como o bíceps braquial), mentres que paralelo pode indicar unha arquitectura lonxitudinal con máis aspecto de fita (como o recto do abdome). Un exemplo menos común son os músculos circulares como o orbicular dos labios, no cal as fibras están dispostas lonxitudinalmente, pero describen un círculo desde a orixe da inserción.
- Arquitectura unipennada
As fibras dos músculos unipennados están todas orientadas co mesmo ángulo relativo con respecto do eixe de xeración de forza (pero non cero).[12] Este ángulo reduce a forza efectiva de calquera fibra individual, xa que efectivamente tira fóra do eixe. Porén, debido a este ángulo, poden empaquetarse máis fibras no mesmo volume muscular, o que incrementa a área transversal fisiolóxica (en inglés, PCSA, Physiological cross-sectional area). Este efecto coñécese como empaquetamento das fibras, e, en termos de xeración de forza, supera sobradamente a eficiencia perdida pola orientación fóra do eixe. A compensación prodúcese na velocidade global de acurtamento do músculo e na excursión total. A velocidade de acurtamento global do músculo é reducida comparada coa velocidade de acurtamento das fibras, xa que é a distancia total de acurtamento.[12] Todos estes efectos dependen do ángulo de pennación; a maiores ángulos maiores forzas xeradas debido ao aumento do empaquetamento das fibras e á área transversal fisiolóxica, pero con grandes perdas na velocidade de acurtamento e excursión do músculo. O músculo vasto lateral é un exemplo de arquitectura unipennada.
- Arquitectura multipennada
As fibras dos músculos multipennados están dispostas en múltiples ángulos en relación co eixe de xeración da forza, e son a arquitectura máis xeral e común.[12] Dentro desta categoría caen varias orientacións das fibras, que se denominan: bipennada, converxente, e multipennada. Aínda que a determinación da área transversal fisiolóxica é máis difícil nestes tipos de arquitectura muscular, aplícase a mesma compensación que se mencionou antes.
As disposicións bipennadas son esencialmente "uves" de fibras amontoadas unhas sobre outras, como as do recto femoral.
As arquitecturas converxentes teñen forma de triángulo ou abano, con orixes musculares largas e insercións máis estreitas.[11] A ampla variación dos ángulos de pennación desta arquitectura pode utilizarse para moitas funcións. Por exemplo, o músculo trapecio, que é un músculo converxente prototípico, contribúe tanto á elevación do ombreiro coma á súa depresión.
As arquitecturas multipennadas non están limitadas a un arranxo das fibras particular, senón que, cando se usa o termo especificamente, se refire xeralmente ao que esencialmente é unha combinación dos tipos bipennado e unipennado con disposición converxente das fibras. Un exemplo desta arquitectura é o músculo deltoide humano.
Fisioloxía celular e contracción
[editar | editar a fonte]Ademais dos compoñentes actina e miosina que constitúen o sarcómero, as fibras musculares esqueléticas tamén conteñen outras dúas protreínas reguladoras importantes, a troponina e a tropomiosina, que son necesarias para a contracción muscular. Estas proteínas están asociadas coa actina e cooperan para impedir que esta interaccione coa miosina. As células do múisculo esquelético son excitables e están suxeitas a despolarización causada polo neurotransmisor acetilcolina, liberado na unión neuromuscular polas neuronas motoras.[13]
Unha vez que a célula foi suficientemente estimulada, o retículo sarcoplásmico da célula libera ións calcio (Ca2+), que despois interaccionan coa proteína reguladora troponina. A troponina unida a calcio experimenta un cambio conformacional que causa o movemento da tropomiosina do seu lugar, o que deixa expostos os sitios de unión para a miosina que ten a actina (antes tapados pola tropomiosina). Isto permite que a miosina acceda a eles e se formen pontes cruzadas entre a actina e a miosina nun ciclo dependente do ATP, e que se acorte o músculo.
Vías de transdución de sinais
[editar | editar a fonte]O fenotipo do tipo de fibra muscular esquelética en animais adultos está regulado por varias vías de sinalización independentes. Estas inclúen vías implicadas na vía Ras/proteína quinase activada por mitóxeno (MAPK), da calcineurina, da proteína quinase dependente do calcio/calmodulina IV, e do coactivador do proliferador do peroxisoma γ 1 (PGC1). A vía de sinlaización Ras/MAPK liga as motoneuronas e os sistemas de sinalización, acoplando a excitación e a regulación da transcrición para promover a indución dependente dos nervios do programa lento en músculos en rexeneración. A calcineurina, unha fosfatase activada polo Ca2+/calmodulina implicada na especificación do tipo de fibra dependente da actividade do nervio no músculo esquelético, controla directamente o estado de fosforilación do factor de transcrición NFAT, o que permite a súa translocación ao núcleo e produce a activación de proteínas musculares de tipo lento en cooperación coa proeína factor potenciador de miocitos 2 (MEF2) e outras proteínas reguladoras. A actividade da proteína quinase dependente de Ca2+/calmodulina está tamén regulada á alza pola actividade de motoneuronas lenta, posiblemente porque amplifica as respostas xeradas pola calcineurina de tipo lento ao promover as funcións do transactivador MEF2 e aumentar a capacidade oxidativa por medio da estimulación da bioxénese mitocondrial.
Os cambios inducidos pola contracción muscular no calcio intracelular ou as especies reactivas do oxíxeno proporcionan sinais a varias vías entre as que están as MAPKs, a calcineurina e a proteína quinase dependente do calcio/calmodulina IV para activar factores de transcrición que regulan a expresión xénica e a actividade encimática no músculo esquelético.

O activador transcricional de receptores nucleares PGC1-α (PPARGC1A), é importante para a regulación de varios xenes mitocondrias implicados no metabolismo oxidativo, e interacciona directamente con MEF2 para activar sinerxicamente os xenes musculares selectivos das fibras de contracción lenta (ST) e tamén serve como diana para a sinalización de calcineurina. Unha vía transcricional mediada polo do receptor activado polo proliferador do peroxisoma δ (PPARδ) está implicado na regulación do fenotipo da fibra muscular esquelética. Os ratos que levan unha forma activada de PPARd mostran un fenotipo de “resistencia”, cun incremento coordinado de encimas oxidativos e de bioxénese mitocondrial e unha maior proporción de fibras de contracción lenta. Desta maneira, por xenómica funcional, a quinase dependente de calcineurina, a quinase dependente de calmodulina, a PGC-1α, e o PPARδ activado forman a base dunha rede de sinalización que controla a transformación do tipo de fibra do músculo esquelético e o perfil metabólico que protexe contra a resistencia á insulina e a obesidade.
A transición desde o metabolismo aeróbico ao anaeróbico durante o esforzo muscular intenso require que varios sistemas se activen rapidamente para asegurar unha subministración constante de ATP para os músculos en funcionamento. Entre estes están un interruptor que cambia de combustibles metabólicos baseados en graxas a outros baseados en carbohidratos, unha redistribución do fluxo sanguíneo desde os músculos en repouso aos músculos en exercicio, e a eliminación de varios dos subprodutos do metabolismo anaeróbico, como o dióxido de carbono e o ácido láctico. Algunhas destas respostas están gobernadas por un control transcricional do fenotipo glicolítico das fibras de contracción rápida (FT). Por exemplo, a reprogramación do músculo esquelético dun fenotipo glicolítico de fibra de contracción lenta a un fenotipo glicolítico de fibra de contracción rápida implica ao complexo Six1/Eya1, composto de membros da familia da proteína Six. Ademais, identificouse o factor 1-α inducible por hipoxia (HIF1A) como un regulador mestre para a expresión de xenes implicados en respostas hipóxicas esenciais que manteñen os niveis de ATP nas células. A ablación xenética do HIF-1α no músculo esquelético foi asociada cun incremento da actividade de encimas limitantes da velocidade na mitocondria, o que indica que o ciclo do ácido cítrico e o aumento da oxidación dos ácidos graxos nestes animais pode estar compensando a diminución do fluxo da vía gicolítica. Porén, as respostas do HIF-1α mediadas pola hipoxia están tamén ligadas á regulación da disfunción mitocondrial por medio da formación excesiva de especies reactivas do oxíxeno na mitocondria.
Hai outras vías que tamén teñen influencia nas característicads do músculo adulto. Por exemplo, a forza física interna dun músculo pode liberar o factor de transcrición factor de resposta sérico (SRF) desde a proteína estrutural titina, o que orixina a alteración do crecemento do músculo.
Investigación
[editar | editar a fonte]Para investigar as propiedades do músculo esquelético utilízanse moitas técnicas. A estimulación muscular eléctrica utilízase para determinar a forza e velocidade de contracción a diferentes frecuencias de estimulación, o que está relacionado coa composición de tipos de fibras e a mestura das mesmas nun determinado grupo muscular. As probas de músculos in vitro utilízanse para unha caracterización máis completa das propiedades do músculo.
A actividade eléctrica asociada coa contracción muscular mídese por electromiografía (EMG). A EMG é unha técnica común utilizada en moitas disciplinas das ciencias do exercicio e a rehabilitación. O músculo esquelético ten dúas respostas fisiolóxicas: relaxación e contracción. Os mecanismos para que teñan lugar xeran unha actividade eléctrica medida por EMG. A EMG pode medir o potencial de acción dun músculo esquelético, o cal se produce por hiperpolarización dos axóns motores a partir dos impulsos nerviosos enviados ao músculo. A EMG utilízase para saber se o músculo estudado está sendo activado, a cantidade de forza xerada, e tamén como un indicador da fatiga muscular.[14] Hai dous tipos de EMG: intramuscular e de superficie (este último máis común). Os sinais da EMG son moito maiores cando un músculo esquelético se contrae que cando se relaxa. Porén, en músculos esqueléticos máis pequenos e profundos os sinais de EMG están reducidos e, por tanto, esta técnica para eles é menos valiosa á hora de medir a súa activación.[15] Nas investigacións nas que se usa a EMG, normalmente realízase a contracción voluntaria máxima no músculo esquelético de interese, para así ter datos de referencia para o resto dos rexistros de EMG que se farán despois durante a proba experimental principal para ese mesmo músculo.[16]
B. K. Pedersen e os seus colegas demostraron que o músculo funciona en parte como un órgano endócrino ao segregar moléculas reguladoras como citocinas e outros péptidos, agora chamados miocinas. As miocinas crese que son mediadores nos beneficios para a saúde que ten o exercicio.[17]
Patoloxías
[editar | editar a fonte]- Artigo principal: Enfermidades neuromusculares.
As doenzas do músculo esquelético denomínanse miopatías, e as doenzas dos nervios denomínanse neuropatías. Ambas as dúas poden afectar á función muscular ou causar dor muscular, polo que con frecuencia se denominan enfermidades neuromusculares.
Notas
[editar | editar a fonte]- ↑ Saladin, Kenneth S. (2010). Anatomy and Physiology (3rd ed.). New York: Watnick. pp. 405–406. ISBN 9780072943689.
- ↑ 2,0 2,1 2,2 2,3 2,4 2,5 MacIntosh, Brian R.; Gardiner, Phillip F.; McComas, Alan J. (2006). Skeletal Muscle: Form and Function. Human Kinetics. ISBN 978-0-7360-4517-9.
- ↑ 3,0 3,1 Smerdu, V; Karsch-Mizrachi, I; Campione, M; Leinwand, L; Schiaffino, S (Dec 1994). "Type IIx myosin heavy chain transcripts are expressed in type IIb fibers of human skeletal muscle.". The American journal of physiology 267 (6 Pt 1): C1723–8. PMID 7545970.
- ↑ Pette, D; Staron, RS (Sep 15, 2000). "Myosin isoforms, muscle fiber types, and transitions.". Microscopy Research and Technique 50 (6): 500–9. PMID 10998639. doi:10.1002/1097-0029(20000915)50:6<500::AID-JEMT7>3.0.CO;2-7.
- ↑ Staron, Robert S.; Johnson, Peter (November 1993). "Myosin polymorphism and differential expression in adult human skeletal muscle". Comparative Biochemistry and Physiology Part B: Comparative Biochemistry 106 (3): 463–475. doi:10.1016/0305-0491(93)90120-T.
- ↑ Buchthal, F; Schmalbruch, H (Aug 1970). "Contraction times and fibre types in intact human muscle.". Acta physiologica Scandinavica 79 (4): 435–52. PMID 5472111. doi:10.1111/j.1748-1716.1970.tb04744.x.
- ↑ Garnett, R A; O'Donovan, M J; Stephens, J A & Taylor, A (1979-02-01). "Motor unit organization of human medial gastrocnemius.". The Journal of Physiology (en inglés) 287 (1): 33–43. PMC 1281479. PMID 430414. doi:10.1113/jphysiol.1979.sp012643. Arquivado dende o orixinal o 12 de setembro de 2019. Consultado o 15 de xullo de 2019.
- ↑ 8,0 8,1 [1] Arquivado 21 de novembro de 2007 en Wayback Machine. Sports Medicine About.com
- ↑ 9,0 9,1 Johnson, M_A, et al. "Data on the distribution of fibre types in thirty-six human muscles: an autopsy study." Journal of the neurological sciences 18.1 (1973): 111-129.
- ↑ Michael Yessis (2006). Build A Better Athlete. Ultimate Athlete Concepts. ISBN 978-1-930546-78-3.
- ↑ 11,0 11,1 11,2 Martini, Frederic H.; Timmons, Michael J.; Tallitsch, Robert B. (2008). Human Anatomy (6 ed.). Benjamin Cummings. pp. 251–252. ISBN 978-0-321-50042-7.
- ↑ 12,0 12,1 12,2 12,3 Lieber, Richard L. Skeletal muscle structure, function, and plasticity. Wolters Kluwer Health, 2002.
- ↑ Costanzo, Linda S. (2002). Physiology (2nd ed.). Philadelphia: Saunders. p. 23. ISBN 0-7216-9549-3.
- ↑ Cè, E; Rampichini, S; Limonta, E; Esposito, F (Dec 10, 2013). "Fatigue effects on the electromechanical delay components during the relaxation phase after isometric contraction.". Acta physiologica (Oxford, England) 211 (1): 82–96. PMID 24319999. doi:10.1111/apha.12212.
- ↑ Xu, Q; Quan, Y; Yang, L; He, J (Jan 2013). "An adaptive algorithm for the determination of the onset and offset of muscle contraction by EMG signal processing.". IEEE transactions on neural systems and rehabilitation engineering : a publication of the IEEE Engineering in Medicine and Biology Society 21 (1): 65–73. PMID 23193462. doi:10.1109/TNSRE.2012.2226916.
- ↑ Milder, DA; Sutherland, EJ; Gandevia, SC; McNulty, PA (2014). "Sustained maximal voluntary contraction produces independent changes in human motor axons and the muscle they innervate.". PLoS ONE 9 (3): e91754. PMC 3951451. PMID 24622330. doi:10.1371/journal.pone.0091754.
- ↑ Pedersen BK. "Muscle as a secretory organ." American Physiological Society. Compr Physiol 3:1337-1362, 2013. http://www.inflammation-metabolism.dk/index.php?pageid=21&pmid=23897689