Článkonožce
| Článkonožce | |
 | |
| Vedecká klasifikácia | |
|---|---|
| Vedecký názov | |
| Arthropoda Gravenhorst, 1843[1] | |
| Vedecká klasifikácia prevažne podľa tohto článku | |
Článkonožce (staršie: článkonohé[2]; lat. Arthropoda) sú najpočetnejší živočíšny kmeň. Je to obrovská, dosť starobylá a vnútorne veľmi diferencovaná skupina článkovaných živočíchov, ktoré fylogeneticky nadväzujú na Polychaeta. V procese fylogenézy však získali znaky, ktoré im umožnili osídliť všetky suchozemské i sladkovodné ekosystémy a vrátiť sa aj naspäť do mora, z ktorého vzišli. Všade, kde sa vyvíjali, dosiahli mimoriadnu mnohorakosť foriem a prekvapujúcu diverzitu (na súši tvoria podľa niektorých odhadov dokonca až 95 % živočíšnych druhov).
Anatómia
[upraviť | upraviť zdroj]Telo sa skladá v najjednoduchších prípadoch (Myriapoda) z hlavy a takmer rovnakých článkov (segmentov), u väčšiny je však segmentácia výrazne heteronómna (články sa od seba líšia stavbou). Stavbu tela a telových príveskov odvodzujeme z hypotetického východiskového modelu, ktorý mal na každom segmente (okrem prvého článku akronu a posledného pygidia) 1 pár článkovaných, s takmer stopercentnou istotou dvojvetvových končatín. Tomuto modelu sú najbližšie trilobity, ale nie sú mu príliš vzdialené ani primitívne kôrovce Crustacea a viacnôžky Myriapoda; druhá skupina však už má končatiny druhotne jednovetvové. Telo vyšších článkonožcov sa primárne člení na tri oddiely: hlavu (caput), hruď (thorax) a bruško (abdomen). Končatiny sú u podstatnej väčšiny vyvinuté len na hrudi (sú to primárne kráčavé nohy), na hlave z nich vznikajú špecializované prívesky, a na brušku zanikajú alebo prekonávajú zložitú zmenu tvaru a funkcie – modifikáciu.
Kutikula
[upraviť | upraviť zdroj]Telo je kryté chitínovou kutikulou, ktorú produkuje epidermis. Kutikula sa skladá z troch častí:
- epikutikula, tenká povrchová pigmentovaná vrstva;
- exokutikula, vrstva chitínu s trámčekovou štruktúrou a systémom jemných vertikálnych kanálikov;
- endokutikula, spodná vrstva so striedavým uložením chitinózno-bielkovinových fibríl (táto štruktúra dosť pripomína preglejku).

Hrúbka a mikroštruktúra týchto vrstiev a niektoré prímesové látky ovplyvňujú konečnú tvrdosť kutikuly – táto môže byť relatívne mäkká (Arachnoidea), ale aj pevná a tvrdá (mnohé skupiny hmyzu). Má viaceré veľmi výhodné vlastnosti – je veľmi odolná voči kyselinám i zásadám, zabraňuje dehydratácii a aj u silne „opancierovaných“ druhov je vždy dostatočne pružná. Má iba jednu nevýhodu a to že zabraňuje v raste. Preto sa článkonožce periodicky zvliekajú – väčšina len v larválnom štádiu, no niektoré po celý život. Proces zvliekania riadi hormón ekdyzón, ktorý produkujú párové predohrudné žľazy (glandulae protothoracales). Kutikula vždy obsahuje istú prímes CaCO3, čo ju robí pevnejšou. Článkonožce s mäkkou kutikulou ho majú prítomný len v stopách, kým u iných skupín pozorujeme silnú (väčšina Diplopoda) alebo veľmi silnú (väčšina Crustacea) inkrustáciu (prestúpenie) CaCO3. Sfarbenie kutikuly je predovšetkým pigmentové (chemické), u mnohých skupín sa však vyskytuje aj sfarbenie fyzikálne – vzniká interferenciou svetla na jemných, ale často zložitých mikroskulptúrach epikutikuly. Týmto spôsobom vznikajú predovšetkým rôzne, často veľmi nápadné kovové lesky hmyzu, zriedka i pavúkov. Pretože kutikula napriek svojej pružnosti umožňuje len veľmi obmedzený pohyb, musel proces sklerotizácie už od začiatku prebiehať paralelne so vznikom segmentácie. Výsledkom týchto fylogeneticky veľmi úspešných procesov je kutikulárny ektoskelet, zložený z mnohých obrúčok a štítkov, ktoré sú spoločne nazývané sklerity. Ektoskelet každého somitu má u fylogeneticky pôvodných typov tvar obrúčky – táto sa skladá z chrbtovej časti (tergitu) a brušnej časti (sternitu), ktoré sú spojené bočným (pleurálnym) blanitým väzom. Takúto stavbu však pozorujeme pomerne zriedka (Myriapoda, časť Crustacea), alebo iba na niektorých častiach tela (abdomen u šesťnožcov Hexapoda). Naopak, u mnohých skupín vznikajú zložité odvodené sklerity (najmä v ektoskelete hrudi).
Hlava
[upraviť | upraviť zdroj]Hlava vzniká zrastom prvých šiestich telových segmentov. Proces vzniku hlavy sa nazýva cefalizácia. Pre aktívne pochopenie vzťahov medzi troma hlavnými recentnými skupinami článkonožcov (Chelicerata, Crustacea a Tracheata) je nevyhnutné venovať hlavovým segmentom a ich príveskom aspoň stručný výklad:

- Segmentum oculare (akron) – je vždy bez príveskov, u všetkých skupín má primárne 1 pár očí, ktoré sú inervované zrakovými lalokmi (lobi optici) z mozgového (cerebrálneho) ganglia.
- Segmentum antennale – u Crustacea a Tracheata má 1 pár jednovetvových tykadiel (antannae). U klepietkavcov Chelicerata je bez príveskov.
- Segmentum intermedium – u Crustacea má 1 pár kratších, ale dvojvetvových tykadielok (antennulae), u klepeitkavcov má 1 pár chelicer, ale u vzdušnicovcov (Tracheata) je bez príveskov, voľný. Prvé 3 hlavové segmenty sú nazývané aj procephalon (neurocephalon); tento obsahuje centrum nervovej sústavy a väčšiu časť receptorov.
- Segmentum mandibulare – u kôrovcov a vzdušnicovcov má 1 pár hryzadiel (mandibulae), u klepietkavcov je voľný.
- Segmentum maxillare – u kôrovcov a vzdušnicovcov majú 1 pár čeľustí (maxillae), u klepietkavcov príslušný pár končatín modifikuje na hmatadlá (pedipalpy). Čeľuste u vzdušnicovcov majú párový prívesok – čeľustné hmatadlá (palpi maxillares), ktoré sú modifikovanými endopoditmi príslušných hlavových končatín.
- Segmentum deuteromaxillare (u vzdušnicovcov segmentum labiale) – u klepietkavcov je voľný, u Crustacea nesie druhý pár čeľustí, ktoré u podstatnej väčšiny Tracheata zrastajú v spodnú peru (labium). Labium má krátky párový prívesok – spodnoperové hmatadlá (palpi labiales), ktorých genéza je podobná ako u čeľustných hmatadiel. Štvrtý až šiesty hlavový segment tvoria deuterocephalon (gnathocephalon), ktorý obsahuje ústne orgány.
Hruď
[upraviť | upraviť zdroj]Hruď (thorax) je u pôvodnejších viacnôžok ešte neodlíšená od bruška, ale už u dvojkrídlovcov sú 3 hrudné somity naznačené tým, že majú len po jednom páre nôh (bruškové somity zrastajú po dva a vytvárajú diplosomity). Zvyšné vzdušnicovce (teda šesťnožce) majú hruď mohutnú, vždy trojčlánkovú. Hruď kôrovcov sa skladá z väčšieho počtu článkov (obyčajne 7 – 8, no niekedy až 11). U primitívnych foriem ju tvoria voľné články, väčšinou však pozorujeme zrastanie jedného, dvoch, alebo všetkých článkov hrudi s hlavou – vzniká tak pereion, nazývaný aj prosoma alebo cephalothorax. U klepietkavcov zrastá hlava s hruďou na pravú hlavohruď (cephalothorax), vzácnou výnimkou sú nohatky (Pantopoda) s voľnými hrudnými článkami Nohatky sa však už v súčasnosti vyčlenujú ako sesterský kmeň klepietkavcov, čiže doň nepatria. U mnohých roztočov (Acarina) ostáva hlava voľná, ale hruď zrastá s bruškom – vzniká tzv. idiosoma, a niekedy sa dokonca všetky 3 telové oddiely spoja do 1 celku (corpus, omniosoma).
Bruško
[upraviť | upraviť zdroj]Bruško (abdomen) má končatiny len u Myriapoda, Merostomata a sčasti u Crustacea, u ostatných pozorujeme úplnú redukciu končatín. Niekedy sú však bruškové končatiny modifikované na snovacie bradavky (čeľaď Araneae) a skoro vždy z nich vzniká pomocný kopulačný aparát samcov.
Končatiny
[upraviť | upraviť zdroj]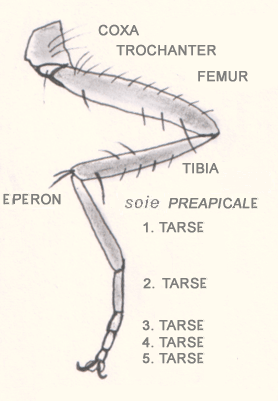
Končatiny sú tiež článkované, primárne dvojvetvové. U väčšiny článkonožcov vonkajšia vetva (exopodit) zaniká, takže končatinu tvorí len vnútorná vetva (endopodit). Niekedy je končatina tiež dvojvetvová, ale jej vetvenie je odvodené, posunuté apikálne (bulbus u pavúkov, klepetá u rakov a niektorých pavúkovcov). Končatina sa primárne skladá z týchto častí:
- coxa (panvička),
- trochanter (predpanvička, niekedy sa používa slovenské označenie „chocholček“),
- femur (stehno),
- patella (koleno),
- tibia (holeň),
- metatarsus (priehlavok) a
- tarsus (chodidlo).
U jednotlivých skupín však pozorujeme množstvo odchyliek – napr. koleno u vzdušnicovcov úplne redukuje, niekedy zaniká i trochanter, a pod. V stavbe nôh a zvlášť modifikovaných končatín sa u článkonožcov prejavuje mimoriadna variabilita.
Svalstvo
[upraviť | upraviť zdroj]Svalstvo článkonožcov je priečne pruhované a upína sa na kutikulárny ektoskelet zvnútra. U viacnôžok je ešte pomerne jednoduché a pripomína fylogeneticky východiskový stav – svalový vak mnohoštetinavcov. U vyšších článkonožcov pozorujeme rozpad svalového vaku v špecializované svalové skupiny – najmä svaly v končatinách sú dosť zložité a ťažko homologizovateľné so svalmi stavovcov (Vertebrata).
Telová dutina
[upraviť | upraviť zdroj]Telová dutina článkonožcov je mixocoel – vzniká splynutím embryonálne zakladaných coelomových dutiniek so zvyškom primárnej telovej dutiny (blastocoelu). Intersegmentálne disepimenty zanikajú, no ich rudimentom zostáva systém lakún – blanitých priepustí medzi sinusmi (dutinkami) otvorenej obehovej sústavy; pričom je zrejmé, že sinusy sú zvyškami coelomových dutiniek. Obehová sústava sa však na rozvode dýchacích plynov podieľa len u kôrovcov a primitívnych pavúkovcov, u vyšších pavúkovcov iba čiastočne (Araneae) alebo vôbec nie (ostatné vyššie Arachnoidea a všetky Tracheata). Pokiaľ hemolymfa plní aj dýchaciu (respiračnú) funkciu, je dýchacím pigmentom hemocyanín, niekedy hemoglobín – časť Crustacea a niektoré vodné larvy (Nematocera). Dýchacie pigmenty sú voľne rozpustené v hemolymfe, nie sú sústredené v erytrocytoch. Hemolymfu uvádza do pohybu trubicovité srdce na chrbtovej strane bruška. Po bokoch má štrbinovité ostie, ktoré majú funkciu chlopní. Pri systole (zmrštení srdca) je hemolymfa vypudená aortou smerom k hlave, potom sa rozlieva v systéme sinusov, až sa dostane opäť do abdomenu, odkiaľ je pri diastole (roztiahnutí srdca) nasávaná do perikardiálneho sinusu a ostiami do srdca. U drobných kôrovcov je niekedy srdce bez aorty (Cladocera), alebo môže úplne chýbať (Ostracoda a väčšina Copepoda). Hemolymfa je vtedy uvádzaná do pohybu peristaltikou čreva a pohybom svalov v tielku.
Dýchacia sústava
[upraviť | upraviť zdroj]Stavba dýchacej sústavy závisí od spôsobu života. Primárne vodné článkonožce (kôrovce a Merostomata) majú žiabre, ktoré sú pôvodne na žiabrových príveskoch (epipoditoch) pri báze hrudných i bruškových končatín; druhotne môžu mať inú polohu – napr. u Decapoda visia do dvoch dutín medzi bočným rozšírením karapaxu a stenou tela. U mnohých drobných druhov žiabre chýbajú a výmena plynov prebieha cez povrchovú jemnú kutikulu (Cladocera, Ostracoda, Copepoda). Dýchacie orgány suchozemských článkonožcov sú dvojakého typu: Pľúcne dutinky a vzdušnice
Pľúcne dutinky
[upraviť | upraviť zdroj]Pľúcne dutinky sú fylogeneticky primitívnejším typom dýchacích orgánov. Sú umiestnené na spodnej strane bruška v počte 1 pár (väčšina Araneae) až 4 páry (Scorpionidea). Vznikli druhotným ponorením žiabrových epipoditov pod úroveň povrchu tela a tu, v uzavretých dutinkách (samozrejme vo vlhku) prebieha výmena plynov. Pľúcne dutinky komunikujú s vonkajškom len cez štrbinovité otvory, ktoré sú chránené viečkom (operculum).
Vzdušnice
[upraviť | upraviť zdroj]Vzdušnice (trachey) sú vývojovo vyšším typom dýchacieho ústrojenstva. Sú to vlastne ektodermálne vliačeniny, spevnené chitinóznou špirálovitou výstužou. Tracheálna sústava vyúsťuje na povrch nenápadnými otvormi (stigmami) po bokoch bruška, niekedy i hrude. Vetvia sa v systém vzdušničiek (tracheol), ktoré privádzajú kyslík priamo k bunkám. Takýto mechanizmus dýchania je veľmi efektívny – vylučuje totiž akékoľvek zdržanie v zásobovaní tkanív kyslíkom, spôsobené u iných skupín živočíchov zložitým prenosom O2 a CO2 dýchacími pigmentami. Znamená to, že svaly Tracheata nikdy nepracujú na kyslíkový dlh, tak častý napr. pri mäkkýšoch, no i inde, vrátane stavovcov. Vzdušnice sú však výhodné len pre telo menších rozmerov. Veľkosť tela vzdušnicovcov má prirodzenú fyziologickú limitu asi 20 – 25 cm, väčšie telo by už tracheálny systém nedokázal efektívne zásobovať kyslíkom. U niektorých lariev vodného hmyzu vznikajú tracheálne žiabre. Sú to vlastne druhotné vyliačeniny kutikuly obyčajne na konci alebo po bokoch bruška, do ktorých vnikajú trachey. K výmene plynov dochádza na jemnej, lamelárne zriasenej kutikule (najády Ephemeroptera, Zygoptera a Plecoptera). Niektoré vodné larvy dýchajú rektálnou dutinou vystlanou tracheolami (Anisoptera), alebo primitívnymi kutikulárnymi žiabrami (Simuliidae). Nakoniec, u suchozemských alebo amfibických článkonožcov platí to isté ako u vodných – u niektorých drobných foriem špecializované dýchacie orgány zanikajú, takže výmena plynov prebieha iba povrchom tela (napr. veľmi drobné roztoče).
Nervová sústava
[upraviť | upraviť zdroj]Nervová sústava je u primitívnejších skupín (viacnôžky, nižšie kôrovce) ešte v podstate rebríčková, väčšinou (u odvodených typov) je už gangliová. Centrom nervovej sústavy je cerebrálne ganglium, alebo mozog. Vzniká zrastom troch párových ganglií neurocephalonu. V hlave je okrem toho aj podhltanová uzlina, ktorá vznikla zrastom troch párov ganglií gnathocephalonu. Inervuje ústne orgány. Z hlavy vedú smerom do tela dva hlavné nervové povrazce, no tieto obyčajne integrujú v jeden povrazec, ktorý v každom somite vytvára uzlinu. Zvlášť veľké sú uzliny v hrudných somitoch, ktoré inervujú zložité svalstvo končatín, a u hmyzu aj lietacie svaly. U odvodených typov článkonožcov pozorujeme časté zrasty ganglií, ktoré korešpondujú s tagmatizáciou (zrastaním hlavných telových oddielov) – napr. u hmyzu s kompaktnou hruďou je len jedna mohutná hrudná uzlina – ganglion thoracale.
Článkonožce majú veľmi rozvinuté zmysly. Receptory sa obyčajne rozdeľujú do troch kategórií:
Mechanoreceptory
[upraviť | upraviť zdroj]Mechanoreceptory reagujú na mechanické podnety: vibrácie, tlak, zemskú gravitáciu, no zrejme i na teplotu a pod. Sú umiestnené na celom povrchu tela, no hlavne na exponovaných častiach: na hlave, končatinách a tykadlách. Sú to tzv. hmatové senzily – poznáme ich v zásade 3 typy:
- pomerne hrubé a tupo zakončené štetiny – setae;
- pri báze hrubšie a ku koncu sa postupne zužujúce chlpovité senzily – chaetae
- jemné a po celej dĺžke veľmi tenké botriotrichie, často nazývané aj etymologiky obráteným termínom trichobotrie.
Posledný typ majú hlavne pavúky a bolo dokázané, že tieto veľmi jemné senzily registrujú aj vibrácie vzduchu – pavúky teda istým spôsobom „počujú“, i keď, samozrejme, na inej úrovni ako cicavce. Diferencovaný sluchový zmysel je u článkonožcov zriedkavý, ale vyskytuje sa. Sú to tzv. tympanálne orgány, ktoré sú vyvinuté u aktívne stridulujúceho hmyzu (Saltatoria, Auchenorrhyncha). Ďalším mechanoreceptorom sú statocysty. U článkonožcov sú vždy v hlave, druhotne môžu splynúť v jeden orgán. U Decapoda sa nachádzajú v bazálnych článkoch prvého páru tykadiel. Zvláštne mechanoreceptory sú i vnútri tela, najmä u hmyzu, a nazývajú sa chordotonálne orgány. Registrujú napätie v jednotlivých častiach tela, odpor prostredia, a tým aj rýchlosť prúdenia vzduchu, vody a pod. Najznámejší je tzv. pedicelový (Johnstonov) orgán, ktorý sa nachádza v druhom článku tykadla ako dôležitý znak triedy Ectognatha. U niektorých skupín (Diptera) môže registrovať aj vibrácie vzduchu, teda zvuk.
Chemoreceptory
[upraviť | upraviť zdroj]Chemoreceptory reagujú na látky rozpustené vo vzduchu a vo vode – predstavujú teda čuchový a chuťový zmysel, u článkonožcov sú tieto vnemy často spojené. Chemoreceptory sú opäť predovšetkým na hlave a sú to dosť jednoduché senzorické jamky alebo papily. Chuťové senzily sú hlavne v ústach a na ústnych príveskoch, čuchové sú najmä na tykadlách. Niektoré skupiny hmyzu, kde samec vyhľadáva samicu na diaľku pomocou feromónov, nemajú v ostrosti čuchu medzi živočíchmi konkurenciu.
Fotoreceptory
[upraviť | upraviť zdroj]
Fotoreceptory registrujú ľudským okom viditeľné, ale často i neviditeľné elektromagnetické žiarenie. Sú to oči niekoľkých typov:
- zložené oči (oculae compositae) – pozostávajú z niekoľko málo až z niekoľko desaťtisíc ommatídií. Prvýkrát sa objavili už u trilobitov, z recentných skupín ich majú Crustacea, Merostomata, Ectognatha, z nadtriedy Myriapoda sa vyskytujú len u Scutigeromorpha;
- ocelli (slovensky označované nepríliš presným termínom „jednoduché očká“) sú v podstate jamkové oči, ktoré pozorujeme u Arachnoidea a Tracheata. U Tracheata (okrem najprimitívnejších) sa vyskytujú len ako akcesorický orgán (hlavným zrakovým orgánom sú zložené oči), ktorý nemá schopnosť obrazového videnia; niekedy sa funkcia ocelli prirovnáva k funkcii expozimetra vo fotoaparáte. U Arachnoidea sú, naopak, hlavným zrakovým orgánom, no ich zrakové schopnosti sú väčšinou slabé. U pavúkov sa však v prednej časti oka vytvorila kutikulárna šošovka, preto ich zrak môže byť už lepší. Relatívne veľmi dobre (a to vo farbách) vidia pavúky, ktoré lovia potravu pomocou zraku (Salticidae, Lycosidae a i.);
- stemmata (ocelli laterales), nazývané niekedy „bočné očká“. Sú to vlastne modifikované ocelli niektorých lariev (húsenice). Majú jednoduchú kutikulárnu šošovku a pod ňou aj kužeľ (conus).
Tráviaca sústava
[upraviť | upraviť zdroj]Tráviaca sústava môže byť rozlične stavaná. Začína sa ústami s ústnymi skleritmi a príveskami, ktoré vznikli modifikáciou hlavových končatín. Nasleduje hltan (pharynx), plynule prechádzajúci do pažeráka (oesophagus), ktorý sa často rozširuje v hrvoľ (ingluvies). Hrvoľ slúži v niektorých prípadoch ako zásobník na predtrávenú alebo rôzne upravenú potravu pre larvy (napr. včely). Tráviaca sústava pokračuje svalnatým alebo žuvacím žalúdkom (proventriculus) a prechádza do žľaznatého žalúdka (ventriculus, mesentheron), ktorý je ako jediný v tráviacej sústave endodermálneho pôvodu. Na rozhraní žuvacieho a žľaznatého žalúdka je často systém slepých výbežkov (coeca), ktoré obsahujú symbiotické mikroorganizmy, u xylofágnych druhov nevyhnutné na trávenie celulózy. Ventriculus má u pôvodných typov (Crustacea, Chelicerata) hepatopankreatickú žľazu, ktorá môže byť tvorená viacerými lalokmi. Okrem sekrečnej funkcie priamo v hepatopankrease prebiehajú tráviace procesy. U drobných foriem (Cladocera, Ostracoda, mnohé Acarina) môže byť hepatopankreas redukovaný. U Tracheata produkuje tráviace enzýmy priamo žľaznatý epitel žalúdka. Žalúdok pokračuje črevom (ileum), ktoré sa končí rektálnou dutinou s análnym otvorom.
Vylučovacia sústava
[upraviť | upraviť zdroj]Vylučovaciu sústavu tvoria orgány dvojakého charakteru:
- Fylogeneticky pôvodnejším typom vylučovacích orgánov sú modifikované metanefrídie. Sú vlastne pozostatkom segmentálne uložených metanefrídií obrúčkavcov Annelida. Pretože telová dutina je už mixocoel, nemusia byť prítomné v každom somite – teoreticky stačí 1 pár, a tak to v skutočnosti aj je. Podľa miesta vyústenia sa nazývajú: 1. koxálne nefrídie (žľazy): vyúsťujú v oblasti panvičiek jedného zo zadných párov končatín (Merostomata, Arachnoidea); 2. maxillárne nefrídie (žľazy): vyúsťujú pri báze druhého páru maxíl (nižšie kôrovce); 3. antennálne nefrídie (žľazy): vyúsťujú v bazálnom článku (scaphus) prvého páru tykadiel (Decapoda).
- Malphigiho trubice (vasa Malphigii) sú charakteristické pre vzdušnicovce, no vyskytujú sa aj u vyšších pavúkovcov (Araneae a Acarina), kde preberajú funkciu hlavného vylučovacieho systému (koxálne nefrídie môžu byť vtedy iba rudimentálne). Malphigiho trubice sú tenkostenné kanáliky, ktoré odvádzajú splodiny metabolizmu z hemolymfy. U vzdušnicovcov majú spoločné vyústenie na začiatku čreva a sú ektodermálneho pôvodu, no u pavúkovcov vyúsťujú do zadnej časti žľaznatého žalúdka a ich pôvod je endodermálny.
S exkrečným systémom súvisí aj tukové teleso (corpus adiposum), prítomné hlavne u lariev. Má primárne zásobnú funkciu, ale hromadia sa v ňom aj niektoré metabolity, ktoré zostali nevylúčené. Nakoniec, k exkrečným mechanizmom patrí aj zvliekanie kutikuly (u lariev, a niekedy i dospelcov). Vo vrstvičke tekutiny, ktorá oddeľuje starú kutikulu od novej, nájdeme tiež určité látky, ktorých sa organizmus potrebuje zbaviť.
Rozmnožovanie a vývin
[upraviť | upraviť zdroj]Sú okrem vzácnych výnimiek (parazitické kôrovce) gonochoristy. Oplodnenie je vnútorné, pomocou penisu alebo modifikovaných končatín. Vajíčko má chitinózny obal, často s charakteristickými skulptúrami. V hornej časti vajíčka je 1 alebo viac otvorov (mikropyle), ktorými sa spermia dostáva k jadru. Väčšina druhov sa vyvíja z oplodneného vajíčka, no pre niektoré skupiny je typická partenogenéza. Poznáme v zásade 3 typy partenogenézy:
- arrhenotokia: samice sa liahnu z oplodnených, kým samce z neoplodnených vajíčok (spoločensky žijúce Hymenoptera). Samce sú haploidné – spermiogenéza u nich prebieha bez meiózy;
- thelytokia: samice sa liahnu z neoplodnených vajíčok (Phasmida, Aphidinea, Otiorhynchus spp., Saga pedo, a i.). U niektorých thelytóknych druhov sa samce vyskytujú len veľmi zriedka alebo vôbec nie, v tom prípade žijú trvalo v samičích populáciách, ktoré sú však diploidné vďaka tzv. náhradnej diploidizácii – vajíčko v procese oogenézy splýva s jednou zo štyroch pólových buniek. (Thelytokia sa vyskytuje aj u iných, primitívnejších živočíšnych skupín, napr. u Rotifera, Chaetonotoidea a pod.);
- amphitokia: z neoplodnených vajíčok vznikajú obe pohlavia. Tento zriedkavý typ partenogenézy nájdeme v životných cykloch vošiek, ďalej u niektorých hrčiarok (Cynipidae) a i.
Vajíčko obsahuje dosť žĺtka a jeho brázdenie je superficiálne, gastrula vzniká epiboliou. Vývin môže byť priamy i nepriamy. Prebieha v mnohých variantoch. Typy premeny sú však veľmi dôležité pre systematické triedenie kmeňa.
Priamy vývin
[upraviť | upraviť zdroj]U Arthropoda, keďže ide o článkované živočíchy, je priamy vývin nazývaný často epiméria; terminologicky jasnejšie by bolo asi označenie juvenilná izoméria. Pri epimérii už 1. instar po vyliahnutí z vajíčka vyzerá ako „zmenšenina“ dospelého živočícha a má hneď plný počet somitov. Vyskytuje sa u väčšiny Arachnoidea, ďalej u Cladocera, Isopoda, Amphipoda, Astacidae, Scolopendromorpha, Geophilomorpha, Thysanura a u triedy Entognatha s výnimkou Protura.
Anaméria
[upraviť | upraviť zdroj]Na hranici priameho a nepriameho vývinu je anaméria alebo juvenilná anizoméria. Pri anamérii juvenilný jedinec vyzerá skoro ako dospelý, líši sa len menším počtom somitov, a niekedy len menším počtom končatín. Definitívny počet somitov a končatín získava medzi jednotlivými zvliekaniami. Typicky anamérne sú napr. Pseudoscorpionidea, Opilionidea, Acarina, všetky Diplopoda, Lithobiomorpha a Protura. U niektorých skupín je na začiatku vývinu larva, ktorá prekonáva premenu na subadultné štádium, ktoré sa mení na dospelého jedinca anamérnym spôsobom (Copepoda, Merostomata, Pantopoda).
Nepriamy vývin
[upraviť | upraviť zdroj]Nepriamy vývin prebieha cez sekundárne larvy; u jednotlivých skupín Arthropoda prichádza do úvahy vlastne 5 variánt:

1. premena väčšiny nižších kôrovcov cez nauplius;
2. premena väčšiny vyšších kôrovcov (Malacostraca) cez zoeu;
3. premena hrotnáčov (Merostomata) cez tzv. trilobitoidnú larvu;
4. premena nohatiek (Pantopoda) cez tzv. phoxichilídium;
V prípadoch 1 až 4 prebieha premena subadultného štádia na dospelého jedinca v podstate anamérnym spôsobom. (Napr. nauplius u Copepoda prekonáva premenu na subadultné, tzv. kopepoditové štádium, ktoré sa mení na dospelého jedinca vlastne anamérnym procesom. Niekedy oddeľuje tieto vývinové štádiá ďalší instar, tzv. metanauplius);
5. premena hmyzu (Ectognatha = Insecta) cez rôzne utvárané sekundárne larvy. Pri hmyze hovoríme viac-menej historicky o dvoch hlavných typoch premeny:
- neúplná premena (hemimetabólia), pri nej vždy chýba pokojové štádium (kukla), pri neometabolii však už môže byť naznačené. Poznáme 4 hlavné typy neúplnej premeny:
- prometabólia: larva žije vo vode a nazýva sa najáda, dýcha tracheálnymi žiabrami. Posledný instar sa zvlieka do štádia subimága, ktoré už má končatiny i mliečne zakalené krídla (a často i lieta), a onedlho sa zvlieka v definitívne imágo (Ephemeroptera);
- archimetabólia: larva – najáda žije tiež vo vode a dýcha tracheálnymi žiabrami (u Anisoptera rektálnou dutinou), chýba však štádium subimága (Plecoptera a Odonata);
- paurometabólia: je to vlastne postupná premena, často blízka anamérii. Larvy sa nazývajú nymfy, žijú spolu s imágami a živia sa tou istou potravou. Podobajú sa imágam, no chýbajú im krídla a pohlavné orgány. Paurometabolia je typická pre celú kohortu Polyneoptera a pre ve1kú väčšinu kohorty Paraneoptera;
- neometabólia: stojí na prechode medzi neúplnou a úplnou premenou. Prvé instary iba kvantitatívne rastú, a premena je posunutá do posledného, príp. posledných dvoch instarov. Obyčajne je naznačené aj kratšie pokojové štádium, pseudopupa. Neometaboliu pozorujeme u Thysanoptera, Aleyrodinea a sčasti u Coccinea (samce);
- úplná premena je charakteristická pokojovým štádiom – kuklou (pupa), v ktorej prebehne úplne celá premena. Znamená to, že larva počas celého larválneho života len kvantitatívne rastie a hromadí zásoby, potrebné na premenu. Poznáme dve varianty úplnej premeny:
- holometabólia: počas premeny je prítomný len 1 typ larvy a jedno pokojové štádium. Takáto premena u hmyzu vysoko prevažuje, vyvíjajú sa ňou takmer všetky druhy kohorty Oligoneoptera (= Holometabola);
- hypermetabólia: zložitý vývin prebieha obyčajne cez 3 rôzne larválne štádiá, prvé je invázne, druhé a niekedy i tretie je parazitické. Jednotlivé larválne štádiá obyčajne oddeľuje krátkodobé pokojové štádium (pseudochrysalis). Hypermetaboliu pozorujeme u Meloidae, Strepsiptera, ale aj inde (chrobáky z čeľade Rhipiphoridae a niektoré parazitoidy zo skupiny Chalcidoidea).
Početnosť
[upraviť | upraviť zdroj]
Dodnes je popísaných asi 1 200 000 druhov článkonožcov, no skutočný počet bude ešte vyšší: minimalistické odhady sú asi 3 milióny, maximalistické až 10 miliónov. Tieto údaje nie sú nadnesené – z asi 80-tich radov článkonožcov sú dôkladne preskúmané vlastne iba motýle a chrobáky, a aj to len vo vyspelejších krajinách sveta. Celé veľké oblasti (sub)trópov, ale i morí ešte len čakajú na systematický výskum väčšiny radov a čeľadí článkonožcov.
Systematika
[upraviť | upraviť zdroj]
Systematika článkonožcov v súčasnosti patrí k najspornejším v celej živočíšnej systematike. O rôznych variantoch systematiky článkonožcov pozri aj články bezhryzadlovce, hryzadlovce a vzdušnicovce.
Systematika primárne používaná v tejto wikipédii je takáto [4][5][6][7][8][9]:
kmeň: článkonožce (Arthropoda)
- oddelenie: bezhryzadlovce (Amandibulata/Arachnomorpha) – existencia tohto oddelenia je dnes sporná
- ...vyhynuté taxóny
- podkmeň † trilobity/trojlalokovce (Trilobitomorpha)
- podkmeň klepietkavce (Chelicerata) – v širšom zmysle, t.j. vrátane nohatiek (Pantopoda)
- oddelenie: hryzadlovce (Mandibulata)
Poznámky:
- Pancrustacea/Tetraconata = Hexapoda + Crustacea
- vzdušnicovce (Tracheata/Atelocerata/Antennata [v užšom zmysle]/Uniramia) = Hexapoda + Myriapoda
- Paradoxopoda/Myriochelata = Chelicerata + Myriapoda
- Antennulata/Antennata [v širšom zmysle] = (takmer) celé článkonožce okrem Chelicerata
Niektorí autori (najmä staršie) do článkonožcov (Arthropoda) zaraďujú aj pomalky (Tardigrada) alebo pazúrikavce (Onychophora) alebo tak pomalky ako aj pazúrikavce. V súčasných systémoch väčšinou Arthropoda nezahŕňajúce pomalky či pazúrikavce nazývame na upresnenie aj Euarthropoda (i keď tento názov sa niekedy používa aj v trochu užších významoch [7]); zoskupenie Euarthropoda+Tardigrada+Onychophora nazývame Panarthropoda (iné názvy: Gnathopoda, Ailopoda, Podophora, Antennopoda); a zoskupenie Euarthropoda+Tardigrada nazývame Tactopoda. [10][11][12][13]
Referencie
[upraviť | upraviť zdroj]- ↑ https://doi.org/10.18476%2F2023.472723
- ↑ MASÁR, I: Niektoré dynamické črty zoologickej a botanickej nomenklatúry. In: Kultúra slova 1985, č. 9, str. 289-294 [1] S. 293
- ↑ Hassanin, Alexandre: Phylogeny of Arthropoda inferred from mitochondrial sequences: Strategies for limiting the misleading effects of multiple changes in pattern and rates of substitution. Molecular Phylogenetics and Evolution 38: 100–116. (2006)
- ↑ PaedDr. Valerián Franc CSc.: Systém a fylogenéza živočíchov – bezchordáty – zdroj, z ktorého (pôvodne) úplne alebo čiastočne čerpal tento článok.
- ↑ TIRJAKOVÁ, Eva; VĎAČNÝ, Peter; KOCIAN, Ľudovít. Systém eukaryotických jednobunkovcov a živočíchov [online]. Bratislava: Katedra zoológie, Prírodovedecká fakulta UK, 2015, [cit. 2018-06-21]. S. 23, 24. Dostupné online.
- ↑ Thanukos, Anna: The Arthropod Story. University of California, Berkeley.
- ↑ a b Mikko's Phylogeny Archive [online]. helsinki.fi, [cit. 2018-04-23]. Dostupné online.
- ↑ Systema Naturae 2000 In: The Taxonomicon & Systema Naturae 2000 [online]. taxonomicon.taxonomy.nl, [cit. 2018-04-23]. Dostupné online.
- ↑ KJER, K. M., SIMON, Ch. et al. Progress, pitfalls and parallel universes: a history of insect phylogenetics. In: Journal of the Royal Society Interface. August 2016, Volume 13, issue 121 (najmä Fig. 5) dostupné online
- ↑ ANDREJEVA-GRIGOROVIČ, A.S. et al. Systematická paleontológia bezstavovcov 1. 2004. dostupné online
- ↑ SIMONETTA, A., DELLE CAVE, L. The phylogeny of the palaeozoic Arthropods. In: Boll. Zool. 47 (suppl.) : 1-19 (1980) dostupné online
- ↑ heslá Arthropoda, Euarthropoda, Gnathopoda, Panarthropoda. In: NOMINA CIRCUMSCRIBENTIA INSECTORUM [online]. insecta.bio.spbu.ru, [cit. 2018-04-23]. Dostupné online.
- ↑ Mikko's Phylogeny Archive [online]. helsinki.fi, [cit. 2018-04-23]. Dostupné online.
Iné projekty
[upraviť | upraviť zdroj]Commons ponúka multimediálne súbory na tému Článkonožce
Wikidruhy ponúkajú informácie na tému Článkonožce
Externé odkazy
[upraviť | upraviť zdroj]- PaedDr. Valerián Franc CSc.: Systém a fylogenéza živočíchov – bezchordáty – zdroj, z ktorého (pôvodne) úplne alebo čiastočne čerpal tento článok.