筋肉

筋肉(きんにく、ラテン語: musculus、ドイツ語: Muskel、英語: muscle)は、動物の持つ組織のひとつで、収縮することにより力を発生させる、代表的な運動器官である[1]。
動物の運動は、主として筋肉によってもたらされる。ただし、細部に於ける繊毛や鞭毛による運動等、若干の例外はある。なお、筋肉が収縮することにより発生する力を筋力と呼び、これは収縮する筋肉の断面積におおよそ比例する。つまり筋力は、筋肉の太さと密接に関係している。
また、食用に供する食肉は主に筋肉であり、脊椎動物の骨格筋は湿重量の約20%をタンパク質が占め[1]、主にこれを栄養として摂取するために食される[2](ただし、食料品店で肉と表示されているものは筋肉だけでなく脂身(脂肪分の塊)も一緒になった状態で、タンパク質ばかりでなく、かなりの高脂肪の状態で販売されていることが多い)。
語源
[編集]英語の「muscle」(マッスル)は、ラテン語で小さなネズミを意味する「musculus」から派生している。これは、筋収縮の様子が皮膚の中でネズミが動く様に見えた事に由来すると考えられる[3][4]。
分類
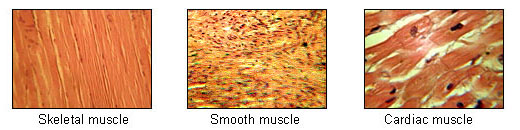
[編集]骨格を持つ動物の筋肉は、その配置から大別すると骨格に付随して身体を構成し、姿勢制御に貢献する骨格筋と、骨格に直接付属せず、身体構成・姿勢制御に直接関わらない内臓筋に分けることができる。しかしこの分類方法は便宜的な分類であり、もっとも良く用いられる分類方法である組織学的分類によれば、多核の骨格筋、単核の平滑筋、心筋に分けることができる[1]。また、意識して動かすことができるかという点で随意筋(骨格筋のみ)と不随意筋(心筋・平滑筋)に分けられる。
- 筋肉
この他にも、筋肉は見た目の色から赤筋(赤色筋) (red muscle, typeI) と白筋(白色筋) (white muscle, typeII) の2種にも分類される。これは含有するミオグロビンやミトコンドリアの量に左右され、多くミトコンドリアが活発なものが赤く、少なく不活発なものが白く見える[5][6] 。またこれらの筋繊維の本数とそれに伴う割合は個人差があり生まれつきほとんど決まっている[要出典]。白筋は収縮の筋原繊維が発達しているため素早く縮むことができるため、速筋 (fast muscle) とも呼ばれる[6]。乳酸性閾値から上の運動強度では速筋が多く使われるようになる[7]。速筋の筋肉繊維は、運動速度や発揮する力によってさらにIIa, IIx, IIbの3種類[8] に分けられる[9]。赤筋は脂肪や炭水化物を消費する酵素が豊富で[9]ゆっくりした運動を持続的に行うのに適し、心臓や呼吸に関する器官の筋肉を構成する[5]。乳酸の代謝では細胞膜を通過して乳酸が輸送される必要がある場合があり、例えば、グリコーゲンが速筋で分解され乳酸を生成し、その乳酸が遅筋や心筋のミトコンドリアで使われている場合がある[7]。一般的な話題で「筋肉」と呼ばれているのは主に骨格筋であり、パワーやスピードの向上に直結するためスポーツでは重要視される。
構造
[編集]解剖学的構造
[編集]
骨格筋
[編集]骨格筋 (skeletal muscle) は、関節など骨格の可動部を動かす筋肉である[10]。脊椎動物では両端が腱を介して骨と繋がった形で配置され、昆虫やエビなどの節足動物ではクチクラ(角皮)を動かすために使われる[10]。関節に関してその筋肉が収縮すると曲がるものを屈筋、伸ばすものを伸筋と言う。その他は回転筋・索引筋・括約筋などに分類される[10]。随意筋であるが、体躯の姿勢制御や反射などでは無意識に動く。体重比で成人男性の42%、同女性の36%を占める[11]。哺乳動物の骨格筋の密度は1.06kg/lであり、脂肪よりも約15%重い[12][13]。
平滑筋
[編集]平滑筋 (smooth muscle) は、横紋が無い筋肉であり、脊椎動物では心臓を除く内臓および血管を構成する筋肉である。無脊椎動物の身体を構成する筋肉はほとんどが平滑筋である[14]。収縮する速度は遅く数十秒かかる場合もあるが、一方で伸び縮みする率は大きく、その状態を保持する能力に優れる[14]。自律神経系から運動の促進・抑制双方の制御を受けている[14]。
心筋
[編集]心筋 (cardiac muscle) は心臓を構成する筋肉である。心筋の特徴として、動作に必要な神経繊維が通常の神経繊維ではなく、特殊心筋と呼ばれる筋群によって興奮が伝達される。従って、肉眼的には神経繊維は存在しない。
微細構造
[編集]筋肉の機能は、神経の制御を受けながら収縮する事と、その収縮度合いを測定しフィードバックすることである。ここでは主な構成要素を、骨格筋を例にして解説し、後に心筋と平滑筋の違いを述べる。
筋繊維(筋線維)/ 筋細胞
[編集]骨格筋を構成する細胞単位。筋芽細胞の融合によって生じる、細長く大きな巨大多核細胞である。骨格筋が発生し分化する過程で、単核の筋原細胞同士が融合してつくられる[15]。
筋原繊維(筋原線維)
[編集]横紋筋の筋繊維中に存在する収縮性の構造体で、細胞内器官。直径約1μmの円筒状をしており、骨格筋では筋肉の長方向に沿って多くの筋原繊維が並行に並んでいる。微細な構造は、多くのサルコメアが厚さ2〜8nmのZ膜(Z線)と呼ばれる隔膜で仕切られながら10nm間隔で連結している。横紋筋の縞模様はこの並びが見えている[16]。ミオフィブリル、筋フィラメント、ミオフィラメントとも呼ばれる[16]。
サルコメア(筋節)
[編集]筋原繊維の最小構成単位。これが縦につながったものが筋原繊維である。個々のサルコメアは、ATP存在下で収縮が起こる。骨格筋の縞は、このサルコメアのアクチンフィラメントとミオシンフィラメントが並行に一部分が重なっている配列に由来する。筋小胞体から放出されたカルシウムイオンにより、アクチンフィラメントがミオシンフィラメントな間に滑り込み筋肉が収縮する。したがって、そのときにはサルコメア全体の長さはアクチンフィラメントが滑り込んだ分だけ小さくなる。
サルコメアには、中央部に密度が高いA帯と、両側に密度が低いI帯がある。A帯は約1.5μm長のミオシンフィラメントで構成され、Z膜に接続したアクチンフィラメントがA帯に入り込んでいない部分がI帯である[16]。両フィラメントは、中心にあるミオシンフィラメントを六角形状にアクチンフィラメントが取り囲んだ断面構造を持つ。ミオシンフィラメント同士の中心間距離は40〜50nm、取り囲むアクチンフィラメントまでの距離は約15nmである[16]。
エネルギー
[編集]筋繊維はアデノシン三リン酸 (ATP) を使い、フィラメント同士がお互い重なり合うように引き付け合い収縮する[1]。
筋肉の制御
[編集]筋肉は、神経からの刺激で収縮を行っている。神経と筋肉は、神経筋接合部というシナプスの一種を介して刺激の伝達を行っている。神経末端からは、アセチルコリンが放出され、筋肉の側にあるアセチルコリン受容体に結合し、筋線維の細胞膜を脱分極させる。これが横行小管(T管)系を伝わって筋全体に広がり、T管系に接する筋小胞体からカルシウムが放出される。このカルシウムをシグナルとして、アクチン繊維とミオシン繊維の間の滑り運動が起こるのである。
その他
[編集]筋繊維は本来積極的に伸展する能力は無く、弛緩したときに伸展するのは、骨格筋の場合、対立筋の働きによる外的な作用による。運動後の筋肉の疲労は、解糖系の最終生成物である乳酸によってもたらされるとの説があるが、医学的根拠は無い。
心筋の微細構造
[編集]心筋は、普通心筋と特殊心筋に分類される、特殊心筋としては、洞房結節、房室結節、ヒス束等が挙げられる。特殊心筋の働きは、心筋の統合された収縮を目的とした、興奮の伝達である。普通心筋は、骨格筋と同じように横紋があるが、骨格筋ほど整然と並んでは居ない。
平滑筋の微細構造
[編集]平滑筋を構成する細胞は紡錘形状で単一の核を持つ[14]。アクチンフィラメントを大量に持ち、ミオシンフィラメントは少量が不規則に分散している。細胞の形状はデスミン中間径フィラメントが存在して保たれる[14]。収縮にはカルシウムイオンによって制御されるが、小胞体があまり発達していないため、細胞膜にあるくびれの外側にイオンを溜め込んでいると考えられる[14]。
筋収縮や弛緩のメカニズム
[編集]大脳に発する運動指令は、小脳において修飾されたのち、遠心性の運動神経を介して、活動電位として伝えられ、運動神経と筋肉の連接部である神経筋接合部に至る。
運動神経の末端にある神経終末(シナプス前末端)に活動電位が伝わると、ここに分布する電位依存性Caチャネルを開口させて、Ca電流を生じる。これによるCa濃度上昇はACh(アセチルコリン)放出を惹起させ、ここで放出されたAChは、シナプス間隙に拡散して、筋肉側で神経終末と結合している終板に達する。終板にはAChのニコチン受容体があり、これにAChが結合することでNa(ナトリウム)、K(カリウム)、Ca(カルシウム)が流入して、いわゆる終板電位 (EPP)を発生させる。これは、筋鞘を介して筋線維全体に伝播されたのち、横行小管 (T管)を介して筋線維の中に入って筋小胞体へ至り、筋小胞体からCa2+の放出を引き起こす。これにより細胞内Ca2+濃度が増加し、トロポニンとCa2+が結合し、トロポニンにアロステリックな変化が生じる。この変化によりトロポミオシンが動き、ミオシンの作用部位が露出する。これによりミオシンとアクチンが反応して相対的な滑りを起こし、筋収縮が引き起こされる[1]。一方、Ca2+は、筋小胞体膜上のCa-ATPaseによって回収され、これによってCa濃度が正常値まで低下するとトロポニンとCa2+の結合が解除され、連鎖的に筋収縮は終了する。
なお、原生動物の組織内にもアクチンやミオシンがフィラメント状に存在している[1]。
起始と停止
[編集]骨格筋は、一般に二つの骨の間にあって、収縮によって骨の位置を変える[17]。骨に付着する筋の両端のうちで、運動時に動きが少ない、又は固定している端を起始(origin)と呼び、大きく動く端を停止(insertion)と呼ぶ[17]。体幹の筋では体幹に近い方が起始、体幹の筋では脊柱に近い方が起始、上下方向の筋では骨盤に近い方が起始と定められている[17]。起始・停止は、一般に結合組織より成る腱となっている[17]。
筋タンパク質
[編集]脊髄動物の骨格筋には、湿潤重量で約20%のタンパク質が含まれ、これを筋タンパク質または筋肉タンパク質という。筋タンパク質の半分は細胞組織である細胞膜・ミトコンドリア・小胞体・細胞核などと、酵素タンパク質が占める。あとの半分は筋原繊維をつくる構造タンパク質であり、アクチン・ミオシンと調整タンパク質・骨格タンパク質などがある[2]。
筋による糖取り込み
[編集]骨格筋は体重の約半分を占める人体最大の器官であり、血糖の80%以上は骨格筋によって取り込まれる。筋では糖輸送担体であるグルコーストランスポーター4型(GLUT4)が糖取り込みの働きを担っている。 GLUT4は普段筋細胞内部に存在しているが、細胞膜へ移動することで、細胞膜上で糖を通過させる門の働きを担う[18]。
インスリンは筋細胞表面に存在する受容体に結合することで、GLUT4を細胞膜へ移動させる働きをする。 また、運動による筋収縮もGLUT4を細胞膜へ移動させる働きがあり、これはインスリンに依存せず血糖の取り込みを亢進する[18]。
運動後の筋も消費された筋グリコーゲンの回復させるために活発に血糖を取り込み続ける必要がある。そのため、運動終了後2~3時間経過した後は、活動筋におけるインスリン感受性が上昇する。 GLUT4タンパク量自体も運動により増加するため、運動・トレーニングを繰り返し行うことにより鍛えられた筋では約2倍近くに増える。これはインスリン刺激による筋の血糖取り込み能力を増加させ、疲労回復を早める効果もある[18]。
発生・発達
[編集]すべての筋肉は沿軸中胚葉から発生している。沿軸中胚葉は胎児の体躯に沿い、体節ごとに分かれている。これは主に3つがあり、脊髄を形成する硬節、皮膚を形成する皮膚分節、筋肉を形成する神経節である。この中で神経節は上下の節に分かれており、それぞれ軸上と軸下の筋肉へとなる。ヒトの場合、上分節は脊柱起立筋と椎間筋肉の一部にしかならない。手足を含むその他の筋肉は全て下分節から発達する[19]。
発生の期間、筋原繊維(筋前駆細胞)は脊椎に関連する筋肉へなるものと、その他の全筋肉を構成するため一度移動して体に取り込まれるものとに分かれる。通常では、側板中胚葉でつくられた筋原繊維がまず外郭を構成する結合組織を作る。そして筋原繊維は化学的な刺激に従いながら、それぞれ適切な場所で骨格筋を形成し始める[19]。
生後、思春期前までは筋肉の発達に男女差が無いが、男性で思春期を迎えると第二次性徴によってアンドロゲンの分泌が活発となり、肩幅が広くなった後に筋肉が発達するようになり[20]、男女間に筋肉の差が生じるようになる。
筋肉は少なくとも2度大きな進化を遂げた。ひとつは刺胞動物であり、もうひとつは左右相称動物である。これらは海綿動物に相当するような有機体にある収縮可能な細胞から進化したものと考えられる[21]。
性差と魅力
[編集]筋肉組織は人間では性的二形である。筋肉組織の性差は上半身で最も明確であり、平均して、男性は女性よりも60%多い総筋肉量と80%多い腕の筋肉を持っている[22]。したがって、男性の上半身の強さは女性よりも約90%高く(ゴリラのオスとメスの違いに匹敵する)[23]、平均的な男性は99.9%の女性よりも強い[24][23]。これらの筋肉質の著しい性差は、攻撃性における強い性差の証拠とともに[25]、人類の進化を通じて男性において物理的対立によって課せられた選択圧力を反映している[26][27]。
選好
[編集]一般的に女性は筋肉質の男性を好むと考えられている。筋肉のある男性は人気のあるアメリカのメディアで魅力的な男性として取り上げられ[28]、他の男性よりも魅力的であるとされ[29][30][31]、より多くの性的なパートナーと経験している[30][32]。また、ほとんどの男性は、女性を引き付け、性的競争でより成功するために、筋肉を増やそうとしている[33]。ただし、非常に筋肉質の男性は、中程度の筋肉質の男性よりも魅力的ではないと評価されている[30]。
男性の筋肉は配偶者として役に立つ情報のシグナリングとして機能していると考えられている。先祖の過去である進化的適応環境(EEA)において、男性の筋肉はその男性の強さに関する正確で直接的な情報を提供して[34]、間接的に戦闘能力を予測させる[35]。より優れた戦闘能力を持った先祖の男性は、他人の資源をよりよく活用し、脅威や搾取、略奪などから自分や仲間の資源を守り、自身の子供を守ることができたと考えられている[27]。また、男性の筋肉は資源を獲得する能力の指標としても重要で、これは文化を超えた男性の配偶者価値の重要な側面でもある[36]。例えば、ハッツァ族の男性の上半身の強さは、男性の配偶者価値の主要な要素である狩猟能力[37]と明確に関連している[38]。同様のことは現代社会でも見られ、戦闘能力と体力は仲間の価値と明確に関連している[39][40]。
さらに、筋肉は男性の全体的な状態の手がかりとして機能している。筋肉組織は、筋肉の成長と維持にエネルギーを割り当てることができる体を必要とするため、筋肉組織は、筋肉を発達させている男性の健康とその基礎となる耐病性に関する間接的で正直な指標である[41]。さらに、筋肉の発達と維持には大量のエネルギー摂取が必要であるため[32]、筋肉質であることは男性がエネルギーを提供する資源(肉など)にアクセスできる能力の指標としても機能する。筋肉は、いくつかの生存関連タスクの成功を高めるのに役立っており、フェイクしづらくコストのかかる質の手がかりとして役立つ要素の一つである[42]。
筋肉組織と筋力によって、女性に潜在的な配偶者に関する重要な情報を提供したことを考えると、現代の女性は男性の体力を正確に評価できると推測され、実際にそうであることが示唆されている[43][34]。こうした評価により、女性は男性の投資と保護の能力を間接的に評価することができる。さらに、体力や筋肉の遺伝率は50%前後であるため[44][45]、女性は男性の筋肉を評価し、自身の子孫に体力や筋肉に関連する遺伝子を提供できる男性を選好しているとも考えられている[42]。
2019年に発表された研究によると、女性が魅力を感じる男性の筋肉部位は、鍛えるのが難しい部位と正に相関している[42]。上位から、腹斜筋、大殿筋、腹筋、上腕二頭筋、肩甲筋、三頭筋、脹脛、三角筋、大腿四頭筋、胸筋となっていた[42]。
数値
[編集]主な動物の筋肉繊維の直径
[編集]数値は David 1977 から[46]。
| 動物 | 筋肉 | 直径(μm) |
|---|---|---|
| キンギョ | 赤筋繊維 | 36.0 |
| (同) | 白筋繊維 | 49.4 |
| アフリカツメガエル | 脚筋 | 169.0 |
| カワラバト | 胸筋 | 26.8 |
| トガリネズミ | 横隔膜 | 18.0 |
| ハツカネズミ | ふくらはぎ(腓筋) | 60.8 |
| ラット | 長指伸筋 | 85.0 |
| (同) | 横隔膜 | 34.0 |
| モルモット | 横隔膜 | 25.0 |
| ネコ | 横隔膜 | 30.0 |
| ブタ | 横隔膜 | 60.0 |
| ヒト | 横隔膜 | 34.0 |
| (同) | 肋間筋 | 50.4 |
| (同) | 三角筋 | 54.2 |
主な動物の筋力
[編集]数値は Prosesser 1973 から[47]。
| 動物 | 筋肉 | 筋力(N/cm2) |
|---|---|---|
| カキ | 貝柱 | 117.7 |
| ラット | 指伸筋 | 29.4 |
| ロブスター | 遅下制筋 | 27.5 |
| ナマケモノ | 横隔膜 | 20.6 |
| カエル | 縫工筋 | 19.6 |
| ネコ | tenuissimus muscle | 13.7 |
| イヌ | 気管平滑筋 | 7.8 |
筋肉の退化
[編集]
哺乳動物において、不活動や飢餓は骨格筋の退化に繋がる。筋肉量の減少は、タンパク質含有量の低下とともに筋肉細胞の減少や矮小化が伴う[48]。
ヒトでは、ベッドにずっと寝ていて運動を長期間行わない状態にあったり、宇宙飛行士が宇宙に滞在したりすると筋肉の弱化や退化が発生する事が知られている。有人宇宙飛行の世界では特に問題視され、無重力空間で過ごした結果、約30%もの筋肉が減ってしまったこともある[49][50]。これはヒトだけではなく、キンイロジリスやオオヒゲコウモリでも同様に起こると報告されている[51]。
年齢とともに、骨格筋の機能や量を維持する能力がゆっくりと減少する現象があり、これは筋肉減少症と呼ばれる。サルコペニアも加齢による筋委縮がみられる。これらの筋委縮病変の原因ははっきりしていないが、骨格筋繊維の維持を助ける「衛星細胞」が徐々に減ってゆく事との関連が疑われ、さらに筋肉量や衛星細胞を維持するために必要な感度もしくは分泌される重要な成長因子の減衰が関係すると考えられ、現在研究がさかんにされている分野である。筋肉減少症は加齢によって一般的に起こり、また実際には病状として扱われないが、高齢者が怪我をしやすくなったり生活に支障を来たしたりする可能性がある[52]。
脚注
[編集]- ^ a b c d e f g 生化学辞典第2版、p.357 【筋肉】
- ^ a b 生化学辞典第2版、p.357 【筋(肉)タンパク質】
- ^ Alfred Carey Carpenter (2007年). “Muscle”. Anatomy Words. October 3, 2012閲覧。
- ^ Douglas Harper (2012年). “Muscle”. Online Etymology Dictionary. October 3, 2012閲覧。
- ^ a b 生化学辞典第2版、p.729 【赤筋】
- ^ a b 生化学辞典第2版、p.995 【白筋】
- ^ a b 八田秀雄、「新たな乳酸の見方」『学術の動向』 11巻 10号 2006年 p.47-50, doi:10.5363/tits.11.10_47
- ^ Larsson, L; Edström, L; Lindegren, B; Gorza, L; Schiaffino, S (July 1991). “MHC composition and enzyme-histochemical and physiological properties of a novel fast-twitch motor unit type”. The American Journal of Physiology 261 (1 pt 1): C93–101. PMID 1858863 2006年6月11日閲覧。.
- ^ a b McCloud, Aaron (30 November 2011). “Build Fast Twitch Muscle Fibers”. Complete Strength Training. 30 November 2011閲覧。
- ^ a b c 生化学辞典第2版、p.496 【骨格筋】
- ^ Marieb, EN; Hoehn, Katja (2010). Human Anatomy & Physiology (8th ed.). San Francisco: Benjamin Cummings. p. 312. ISBN 978-0-8053-9569-3.
- ^ Urbancheka, M; Pickenb, E; Kaliainenc, L; Kuzon, W (2001). “Specific Force Deficit in Skeletal Muscles of Old Rats Is Partially Explained by the Existence of Denervated Muscle Fibers”. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences 56 (5): B191–B197. doi:10.1093/gerona/56.5.B191.
- ^ Farvid, MS; Ng, TW; Chan, DC; Barrett, PH; Watts, GF (2005). “Association of adiponectin and resistin with adipose tissue compartments, insulin resistance and dyslipidaemia”. Diabetes, obesity & metabolism 7 (4): 406–13. doi:10.1111/j.1463-1326.2004.00410.x. PMID 15955127.
- ^ a b c d e f 生化学辞典第2版、p.1187 【平滑筋】
- ^ 生化学辞典第2版、p.355 【筋繊維】
- ^ a b c d 生化学辞典第2版、p.351 【筋原繊維】
- ^ a b c d 伊藤隆『解剖学講義』南山堂、1983年10月5日、15頁。ISBN 978-4-525-10051-3。
- ^ a b c 川中 健太郎「運動と骨格筋GLUT4」『学術の動向』第11巻第10号、公益財団法人日本学術協力財団、2006年10月1日、42-46頁、doi:10.5363/tits.11.10_42、ISSN 1884-7080、2023年10月16日閲覧。
- ^ a b MacIntosh, BR; Gardiner, PF; McComas, AJ (2006). “1. Muscle Architecture and Muscle Fiber Anatomy”. Skeletal Muscle: Form and Function (2nd ed.). Champaign, IL: Human Kinetics. pp. 3–21. ISBN 0-7360-4517-1
- ^ お母さんの基礎知識(思春期・男の子編)(もっと詳しく…)-神奈川県ホームページ[リンク切れ]
- ^ Steinmetz, Patrick R. H.; Kraus, Johanna E. M.; Larroux, Claire; Hammel, Jörg U.; Amon-Hassenzahl, Annette; Houliston, Evelyn; Wörheide, Gert; Nickel, Michael et al. (2012). “Independent evolution of striated muscles in cnidarians and bilaterians”. Nature 487 (7406): 231–234. doi:10.1038/nature11180. ISSN 0028-0836.
- ^ Abe, T (2003-10-01). “Sex differences in whole body skeletal muscle mass measured by magnetic resonance imaging and its distribution in young Japanese adults”. British Journal of Sports Medicine 37 (5): 436–440. doi:10.1136/bjsm.37.5.436. ISSN 0306-3674.
- ^ a b Lassek, William D.; Gaulin, Steven J.C. (2009-09). “Costs and benefits of fat-free muscle mass in men: relationship to mating success, dietary requirements, and native immunity”. Evolution and Human Behavior 30 (5): 322–328. doi:10.1016/j.evolhumbehav.2009.04.002. ISSN 1090-5138.
- ^ Hagen, Edward H.; Rosenström, Tom (2016-02-15). “Explaining the sex difference in depression with a unified bargaining model of anger and depression”. Evolution, Medicine, and Public Health: eow006. doi:10.1093/emph/eow006. ISSN 2050-6201.
- ^ Archer, John (2009-08). “Does sexual selection explain human sex differences in aggression?”. Behavioral and Brain Sciences 32 (3-4): 249–266. doi:10.1017/s0140525x09990951. ISSN 0140-525X.
- ^ Puts, David A. (2010-05). “Beauty and the beast: mechanisms of sexual selection in humans”. Evolution and Human Behavior 31 (3): 157–175. doi:10.1016/j.evolhumbehav.2010.02.005. ISSN 1090-5138.
- ^ a b Sell, Aaron; Hone, Liana S. E.; Pound, Nicholas (2012-03). “The Importance of Physical Strength to Human Males”. Human Nature 23 (1): 30–44. doi:10.1007/s12110-012-9131-2. ISSN 1045-6767.
- ^ Frederick, David A.; Fessler, Daniel M.T.; Haselton, Martie G. (2005-03). “Do representations of male muscularity differ in men's and women's magazines?”. Body Image 2 (1): 81–86. doi:10.1016/j.bodyim.2004.12.002. ISSN 1740-1445.
- ^ Dixson, Barnaby J.; Dixson, Alan F.; Bishop, Phil J.; Parish, Amy (2009-01-13). “Human Physique and Sexual Attractiveness in Men and Women: A New Zealand–U.S. Comparative Study”. Archives of Sexual Behavior 39 (3): 798–806. doi:10.1007/s10508-008-9441-y. ISSN 0004-0002.
- ^ a b c Frederick, David A.; Haselton, Martie G. (2007-06-15). “Why Is Muscularity Sexy? Tests of the Fitness Indicator Hypothesis”. Personality and Social Psychology Bulletin 33 (8): 1167–1183. doi:10.1177/0146167207303022. ISSN 0146-1672.
- ^ Gray, Peter B.; Frederick, David A. (2012-07). “Body Image and Body Type Preferences in St. Kitts, Caribbean: A Cross-Cultural Comparison with U.S. Samples regarding Attitudes towards Muscularity, Body Fat, and Breast Size”. Evolutionary Psychology 10 (3): 147470491201000. doi:10.1177/147470491201000319. ISSN 1474-7049.
- ^ a b Lassek, William D.; Gaulin, Steven J.C. (2009-09). “Costs and benefits of fat-free muscle mass in men: relationship to mating success, dietary requirements, and native immunity”. Evolution and Human Behavior 30 (5): 322–328. doi:10.1016/j.evolhumbehav.2009.04.002. ISSN 1090-5138.
- ^ Frederick, David A.; Buchanan, Gregory M.; Sadehgi-Azar, Leila; Peplau, Letitia Anne; Haselton, Martie G.; Berezovskaya, Anna; Lipinski, Ryan E. (2007-04). “Desiring the muscular ideal: Men's body satisfaction in the United States, Ukraine, and Ghana.”. Psychology of Men & Masculinity 8 (2): 103–117. doi:10.1037/1524-9220.8.2.103. ISSN 1939-151X.
- ^ a b Sell, Aaron; Cosmides, Leda; Tooby, John; Sznycer, Daniel; von Rueden, Christopher; Gurven, Michael (2008-10-21). “Human adaptations for the visual assessment of strength and fighting ability from the body and face”. Proceedings of the Royal Society B: Biological Sciences 276 (1656): 575–584. doi:10.1098/rspb.2008.1177. ISSN 0962-8452.
- ^ Muñoz-Reyes, José Antonio; Gil-Burmann, Carlos; Fink, Bernhard; Turiegano, Enrique (2012-04-18). “Physical strength, fighting ability, and aggressiveness in adolescents”. American Journal of Human Biology 24 (5): 611–617. doi:10.1002/ajhb.22281. ISSN 1042-0533.
- ^ Buss, David M. (1989-03). “Sex differences in human mate preferences: Evolutionary hypotheses tested in 37 cultures”. Behavioral and Brain Sciences 12 (1): 1–14. doi:10.1017/s0140525x00023992. ISSN 0140-525X.
- ^ Apicella, Coren Lee (2014-11). “Upper-body strength predicts hunting reputation and reproductive success in Hadza hunter–gatherers”. Evolution and Human Behavior 35 (6): 508–518. doi:10.1016/j.evolhumbehav.2014.07.001. ISSN 1090-5138.
- ^ Marlowe, Frank W. (2004-12). “Mate preferences among Hadza hunter-gatherers”. Human Nature 15 (4): 365–376. doi:10.1007/s12110-004-1014-8. ISSN 1045-6767.
- ^ Archer, John; Thanzami, Vanlal (2009-09). “The relation between mate value, entitlement, physical aggression, size and strength among a sample of young Indian men”. Evolution and Human Behavior 30 (5): 315–321. doi:10.1016/j.evolhumbehav.2009.03.003. ISSN 1090-5138.
- ^ Muñoz-Reyes, José Antonio; Fernández, Ana María; Flores-Prado, Luis; Guerra, Rómulo; Turiégano, Enrique (2015-07). “Fighting ability influences mate value in late adolescent men”. Personality and Individual Differences 80: 46–50. doi:10.1016/j.paid.2015.02.026. ISSN 0191-8869.
- ^ Kaplan, Hillard S.; Gangestad, Steven W. (2015-09-08). The Handbook of Evolutionary Psychology. Hoboken, NJ, USA: John Wiley & Sons, Inc.. pp. 68–95. ISBN 978-0-470-93937-6
- ^ a b c d Durkee, Patrick K.; Polo, Pablo; Muñoz-Reyes, José Antonio; Rodríguez-Ruiz, Claudia; Losada-Pérez, María; Fernández-Martínez, Ana B.; Turiégano, Enrique; Buss, David M. et al. (2019-04-01). “Men’s Bodily Attractiveness: Muscles as Fitness Indicators” (英語). Evolutionary Psychology 17 (2): 1474704919852918. doi:10.1177/1474704919852918. ISSN 1474-7049.
- ^ Durkee, Patrick K.; Goetz, Aaron T.; Lukaszewski, Aaron W. (2018-03). “Formidability assessment mechanisms: Examining their speed and automaticity”. Evolution and Human Behavior 39 (2): 170–178. doi:10.1016/j.evolhumbehav.2017.12.006. ISSN 1090-5138.
- ^ Silventoinen, Karri; Magnusson, Patrik K. E.; Tynelius, Per; Kaprio, Jaakko; Rasmussen, Finn (2008-05). “Heritability of body size and muscle strength in young adulthood: a study of one million Swedish men”. Genetic Epidemiology 32 (4): 341–349. doi:10.1002/gepi.20308. ISSN 0741-0395.
- ^ THOMIS, MARTINE A. I.; BEUNEN, GASTON P.; MAES, HERMINE H.; BLIMKIE, CAMERON J.; VAN LEEMPUTTE, MARC; CLAESSENS, ALBRECHT L.; MARCHAL, GUY; WILLEMS, EUSTACHIUS et al. (1998-05). “Strength training: importance of genetic factors”. Medicine & Science in Sports & Exercise 30 (5): 724–731. doi:10.1097/00005768-199805000-00013. ISSN 0195-9131.
- ^ R.Flindt 著、浜本哲郎 訳『数値で見る生物学』ジュプリンガー・ジャパン、2007年、31頁。ISBN 978-4-431-10014-0。
- ^ R.Flindt 著、浜本哲郎 訳『数値で見る生物学』ジュプリンガー・ジャパン、2007年、33頁。ISBN 978-4-431-10014-0。
- ^ Fuster, G; Busquets, S; Almendro, V; López-Soriano, FJ; Argilés, JM (2007). “Antiproteolytic effects of plasma from hibernating bears: a new approach for muscle wasting therapy?”. Clin Nutr 26 (5): 658–61. doi:10.1016/j.clnu.2007.07.003. PMID 17904252.
- ^ Roy, RR; Baldwin, KM; Edgerton, VR (1996). “Response of the neuromuscular unit to spaceflight: What has been learned from the rat model”. Exerc. Sport Sci. Rev. 24: 399–425. PMID 8744257.
- ^ “NASA Muscle Atrophy Research (MARES) Website”. 2013年2月2日閲覧。
- ^ Lohuis, TD; Harlow, HJ; Beck, TD (2007). “Hibernating black bears (Ursus americanus) experience skeletal muscle protein balance during winter anorexia”. Comp. Biochem. Physiol. B, Biochem. Mol. Biol. 147 (1): 20–28. doi:10.1016/j.cbpb.2006.12.020. PMID 17307375.
- ^ Roche, Alex F. (1994). “Sarcopenia: A critical review of its measurements and health-related significance in the middle-aged and elderly”. American Journal of Human Biology 6: 33. doi:10.1002/ajhb.1310060107.
参考文献
[編集]- 『生化学辞典第2版』(第2版第6刷)東京化学同人、1995年。ISBN 4-8079-0340-3。
関連項目
[編集]外部リンク
[編集]| 国立図書館 | |
|---|---|
| その他 | |
この項目は、生物学に関連した書きかけの項目です。この項目を加筆・訂正などしてくださる協力者を求めています(プロジェクト:生命科学/Portal:生物学)。 |