Axon
Axon je výběžek nervové buňky (neuronu) sloužící jako informační výstup při přenosu informace mezi neurony nebo mezi neuronem a jinými buňkami (např. buňkami svalu). Z nervové buňky vystupuje typicky jeden axon, který se na svém konci bohatě větví. Axony se dělí na myelinizované a nemyelinizované. Myelinová pochva, která obaluje myelinizované axony je tvořena gliovými buňkami a zvyšuje rychlost a účinnost vedení vzruchu.
Axon popsal poprvé v 19. století německý anatom Otto Friedrich Karl Deiters. Funkční roli axonu v přenosu vzruchů poprvé objasnil španělský anatom Santiago Ramón y Cajal.[1]
Struktura
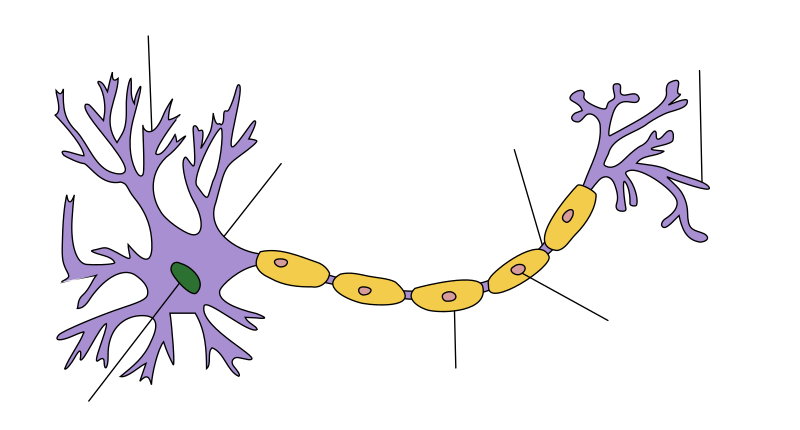
editovatAxon je výběžek neuronu o délce až 100 cm a tloušťce 0,05-20 μm.[2] Axony hlavonožců však mohou dosahovat průměru až 1 mm.[1] Buněčná membrána axonu se nazývá axolemma a cytoplazma axonu axoplazma. Každý neuron má typicky jeden axon a několik bohatě větvených dendritů. U pseudounipolárních buněk odstupuje axon z výběžku společného s dendritem.[3] Axon se dělí na několik částí – nejblíže buněčnému tělu je tzv. odstupový konus (někdy označovaný jako axonový hrbolek), který je následován iniciálním segmentem, což je oblast před počátkem myelinové pochvy. Iniciální segment je místem vzniku akčních potenciálů. V průběhu axonu může docházet k jeho větvení, tyto větve se nazývají kolaterály. Nejdále od buněčného těla se nachází distální konec, kde se axon většinou několikanásobně větví – vytváří terminální arborizaci (telodendron). Jednotlivé větve se na koncích rozšiřují v terminální (synaptické) butony (též označované jako synaptické knoflíky). Synaptické butony však vznikají také podél tenkých axonálních větví, kde utváří ztluštěniny nazývané varikosity.[1][3]
Cytoskelet a buněčné organely
editovatV axonech se hojně vyskytují mikrotubuly (označované jako neurotubuly) a neurofilamenta. Aktinový cytoskelet je v axonu uspořádaný zvláštním způsobem (PMS – periodic membrane skeleton). Aktin v axonu tvoří kroužky pod membránou axonu v pravidelných rozestupech (180-190 nm). Aktinová vlákna jsou stabilizována adducinem, který kontroluje průměr aktinových kroužků. Kroužky jsou propojeny mezi sebou pomocí tetramerů spektrinu. Kroužky vznikají nejprve v části axonu, která je blíže u buněčného těla a během vývoje se poté dostávají i do vzdálených částí axonu. Význam tohoto speciálního uspořádání aktinového cytoskeletu je pravděpodobně mimo vlastního vyztužení a podpory plazmatické membrány také udržování pravidelného rozmístění membránových proteinů důležitých pro udržení struktury axonu a pro přenos vzruchu (např. ankyrin a sodíkové kanály).[4]
V axoplazmě se nachází malé množství mitochondrií a ojediněle také hladké endoplazmatické retikulum. V místech synaptických butonů se nachází mitochondrie a četné synaptické vezikuly (váčky), které obsahují chemické mediátory nezbytné pro přenos vzruchu.[3]
Synapse
editovatNa koncích axonů v místech terminálních butonů nebo v místech varikosit se nachází synapse. Synapse (neboli zápoj) je místem přenosu signálu mezi neurony či mezi neuronem a efektorovou tkání. Každý neuron má řádově tisíce synapsí. Přenosu informace na synapsi říkáme synaptická transmise (neurotransmise). Synapse se dělí na elektrické a chemické. V elektrických synapsích jsou buněčné membrány buněk v těsné blízkosti a jsou vzájemně propojeny membránovými strukturami tvořícími tzv. vodivé spoje (gap junctions tvořené proteiny konexiny), které umožní tok iontů z jedné buňky do druhé. Elektrické synapse jsou rychlejší než chemické a umožňují přenos signálu oběma směry. V lidském nervovém systému se elektrické synapse neuplatňují, najdeme je však například u ryb, obojživelníků či plazů. Chemické synapse přenášejí informace pomocí molekul – neuropřenašečů (neuromediátorů).[5]
Synapse se dělí dle buněk, které se jich účastní, na interneuronové, neuroreceptorové a neuroefektorové. Nejčastější jsou interneuronové synapse, při kterých přechází signál od presynaptického neuronu k postsynaptickému neuronu. Nejčastějším typem synapse mezi neurony je synapse axo-dendritická, dalším typem je synapse axo-somatická a nejméně časté jsou synapse axo-axonální. Axony tvoří synapse pouze v nemyelinizovaných místech. Nejvýznamnějším typem neuroefektorové synapse je nervosvalová ploténka, kde zakončení axonu kontaktuje vlákno příčně pruhovaného svalu.[5]
Z hlediska stavby se synapse skládá z presynaptické části, synaptické štěrbiny a postsynaptické části. Presynaptická část je nejčastěji knoflíkovitě rozšířená koncová část axonu, která obsahuje neurotubuly, neurofilamenta, mitochondrie a četné synaptické vezikuly naplněné neuropřenašečem. Synaptická štěrbina je prostor mezi membránami buněk tvořících synapsi, je široký asi 30–40 nm. Postsynaptická část obsahuje ve své membráně zejména četné receptory pro neuropřenašeče.[5]
Synapse jsou vysoce dynamické útvary a jejich dynamické změny představují synaptickou plasticitu, což je schopnost modifikovat stavbu a činnost existujících synapsí a je to jedna z hlavních možností učení a vytváření paměti.[5]
Myelinizace
editovatMyelinizace (neboli obalení axonů myelinovou pochvou) umožňuje rychlé a účinné vedení vzruchu a představuje tak evoluční výhodu umožňující rychlejší reakce na okolní prostředí. Existují dvě hlavní strategie pro zrychlení vedení vzruchů – zvětšení neuronů a průměru jejich axonu (u bezobratlých, např. chobotnice) a nebo myelinizace, která umožňuje rychlé saltatorní (skokové) vedení. Zvětšení průměru axonu až na 1 mm umožní rychlost vedení až kolem 25 m/s, tato varianta je nicméně prostorově i energeticky velmi náročná. Myelinizace umožňuje rychlost vedení vzruchů kolem 100 m/s.[6]
Myelinová pochva je lipoproteinový komplex (asi 70 % tvoří lipidy, zejména galaktosfingolipidy, sfingomyelin, cholesterol) složený z řady koncentrických vrstev, které vznikají mnohočetným spirálním obtáčením plazmatické membrány gliové buňky kolem úseku axonu.[6]
V periferní nervové soustavě (PNS) je myelinová pochva tvořena Schwannovými buňkami, kde každá Schwannova buňka vytvoří jeden myelinový plát kolem jednoho axonu. V centrální nervové soustavě (CNS) je myelinová pochva produktem oligodendrocytů, které jsou schopny vytvořit více myelinových pochev na různých axonech (1 oligodendrocyt tvoří kolem 30 myelinových pochev). Myelin na jednom axonu je tedy tvořen několika myelinovými pláty umístěnými podél axonu a každý tento plát je tvořen jinými oligodedrocyty či Schwannovými buňkami. Mezi těmito segmenty se nachází tzv. Ranvierův zářez (nodus). V této oblasti se akumulují napěťově řízené sodíkové kanály, což spolu s izolačními vlastnostmi myelinu a jeho spojení s membránou axonu, vede k saltatornímu (skokovému) vedení vzruchu.[6]
Ne všechny axony obratlovců jsou však myelinizovány. Např. v periferní nervové soustavě nemají myelinovou pochvu post-ganglionická autonomní vlákna nebo některá vlákna pro cítění bolesti a teploty. V CNS zůstávají bez myelinizace např. paralelní vlákna v mozečku. Také nejtenčí axony nemají myelinovou pochvu a označují se jako nemyelinizovaná či šedá vlákna, která jsou v PNS obalena cytoplazmou Schwannovy buňky, zatímco v CNS zůstávají tato vlákna holá a jsou součástí tzv. neuropilu.[6][3]
Myelinizace vede k organizaci axonálních proteinů do specifických domén a vedle urychlení vedení vzruchu hraje roli také v neuronální plasticitě. Myelinová pochva má význam také pro energetické potřeby axonu, které spočívají zejména v axonálním transportu na dlouhé vzdálenosti. Gliové buňky tvořící myelinovou pochvu jsou zásadní pro zásobení axonů energetickými metabolity (zejména laktátem).[6]
Nervy
editovatAxony (neboli nervová vlákna) se sdružují ve větší útvary nazývané nervy. Axony obalené myelinovou pochvou jsou obklopeny tenkou vrstvou jemného vaziva nazývanou endoneurium. Svazek takových axonů (neboli fascikulus) je obalen plochými buňkami perineuria a několik takových svazků obklopených společně fibrózní vrstvou pojivové tkáně (epineurium) utváří periferní nerv.[7]
Dle směru, kterým nervová vlákna přenáší informaci je dělíme na aferentní neboli sensitivní (informace od periferie do centra – centripetálně) a na eferentní (motorická vlákna k efektorovým orgánům). Kontakt obou druhů těchto vláken vytváří reflexní oblouk.[8]
Vývoj a regenerace
editovatVývoj axonu
editovatNeurony vznikají diferenciací z neurálních progenitorů, které vznikají z neurálních kmenových buněk. Neurální progenitor je oválná buňka bez výrazných výběžků. Během diferenciace začnou neurony vytvářet několik tenkých výběžků zvaných neurity. Tento děj je významně ovlivňován buněčným cytoskeletem zejména mikrotubuly a aktinovými vlákny. Svazky mikrotubulů tvoří kostru vznikajících výběžků a pravděpodobně se účastní vzniku výběžků díky takzvanému “klouzání“ mikrotubulů, kdy nedochází k prodlužování stávajícího mikrotubulového vlákna, ale spíše k posouvání mikrotubulů vůči sobě, čehož se účastní molekulární motory (proteiny schopné pohybu po mikrotubulech). Posun mikrotubulového vlákna vyvíjející tlak na buněčnou membránu může spolu s přestavbami aktinového cytoskeletu pod membránou vést ke vzniku neuritu. Jeden ze vzniklých výběžků se následně stane axonem, zatímco zbývající výběžky budou utvářet dendrity. Ve výběru budoucího axonu hraje významnou roli opět cytoskelet a to zejména jeho stabilita. Výběžek s vysokým podílem stabilizovaných mikrotubulových vláken a zároveň nestabilními dynamickými aktinovými vlákny utvoří budoucí axon. Jedno z vysvětlení tohoto procesu je, že stabilizovaný mikrotubul je selektivně využíván některými molekulárními motory se specifickým nákladem, a tedy ovlivní buněčný transport, což může vést k hromadění organel a proteinů nezbytných pro vznik axonu právě ve výběžku s nejstabilnějšími mikrotubuly. Tohoto selektivního transportu se patrně účastní Kinesin-1, který se váže především na stabilní mikrotubuly a bylo popsáno, že se hromadí v budoucím axonu.[9]
Ve chvíli, kdy vzniká axon, dochází k polarizaci neuronu. Během tohoto procesu dochází k zániku MTOC (mikrotubuly organizující centrum), z kterého mikrotubuly vychází ve většině ostatních buněk. V neuronech jsou minus konce mikrotubulů stabilizovány např. na membráně Golgiho aparátu nebo pomocí proteinu CAMSAP2 (calmodulin-regulated spectrin-associated protein 2), jehož přítomnost je potřebná pro vznik axonu.[9]
Růstový kužel a navádění axonů
editovatPo utvoření axonu dochází ke zvětšení jeho koncové části a utvoření růstového kužele, což je dynamická struktura na konci axonu zodpovědná za růst a větvení axonu. Růstový kužel také na základě chemických (např. semaforiny) a fyzikálních podnětů z okolí řídí růst axonu do správného směru.
Růstový kužel se skládá z periferní domény, která je tvořena lamelipodii a filopodii vyplněnými aktinem (aktinové svazky utváří filopodia a větvené aktinové sítě lamelipodia). Centrální doména obsahuje mikrotubuly směrované svým dynamickým rostoucím koncem k periferní doméně. Část axonu za růstovým kuželem směrem k tělu neuronu se označuje jako axonální shaft. V této oblasti jsou mikrotubuly více stabilizovány v porovnání s dynamickým růstovým kuželem. Této stability je dosaženo pomocí posttranslačních modifikací tubulinu tvořícího mikrotubuly (detyrosinace, acetylace).[9]
Růst axonů se rozděluje do 3 fází – v první fázi dochází k protahování filopodií a lamelipodií způsobenému polymerizací aktinu. Vlákna aktinu polymerující ve směru růstu zároveň depolymerují na opačné straně, čímž vznikne prostor pro polymerující mikrotubuly (fáze 2). Nově polymerované mikrotubuly jsou ve třetí fázi stabilizovány, čímž se utváří nový axonální shaft.[9]
Schopnost růstu axonu je omezena po vzniku synapse a jeho zapojení do neuronové sítě. Po narození připadá na jeden neuron mozkové kůry cca 2500 synapsí a ve věku kolem 3 let asi 15 000 synapsí. U dospělého člověka je průměrný počet synapsí nižší (cca poloviční), neboť během vývoje dochází k zániku nevyužívaných synapsí, tomuto jevu se říká synaptické okleštění (synaptic pruning).[10]
Regenerace
editovatV periferní nervové soustavě si neurony uchovávají schopnost reaktivace růstu a tedy regenerace. Nicméně při výrazném poškození nemusí být regenerace dostatečná, nedojde k opětovné inervaci a může dojít ke vzniku chronické bolesti.[9]
V centrální nervové soustavě je regenerace možná pouze během embryonálního vývoje. Po poškození axonů v dospělé CNS dochází k přestavbě cytoskeletu v místě přerušení axonu, které následně brání regeneraci. Následkem toho nejsou poškozené axony v CNS schopné vytvořit růstový kužel a na konci axonu utvoří oválné struktury (retraction bulb), které se časem zvětšují. Namísto uspořádání cytoskeletu typického pro růstový kužel, kde jsou mikrotubuly spíše v centrální části a mikrofilamenta na povrchu, se po poškození axonu v CNS tyto cytoskeletární struktury překrývají. To je příčinou nevratných poškození po mrtvici či poškození míchy. Regeneraci axonů v CNS brání také inhibiční molekuly sekretované oligodendrocyty a přítomnost buněk tvořících jizvu.[11][9]
Funkce
editovatAxon slouží k přenosu informace pomocí vedení vzruchů. V iniciálním segmentu axonů se vstupy z buněčného těla přetváří v akční potenciál, který se následně šíří podél axonu k jeho zakončení rychlostí asi 100 m/s. Rychlost vedení vzruchu je ovlivněna průměrem vlákna a myelinizací.[12]
Membránový potenciál
editovatNa membráně všech živých buněk existuje elektrické napětí mezi vnitřním prostředím buňky a okolím dané nerovnoměrným rozdělením iontů uvnitř a vně buňky. Tomuto napětí se říká klidový membránový potenciál. Při klidovém membránovém potenciálu je koncentrace K+ iontů v buňce asi 10× vyšší než koncentrace v mezibuněčném prostoru. Naopak koncentrace Na+ a Cl− iontů je výrazně vyšší vně buňky. Tento nepoměr je udržován pomocí Na+/K+ ATPázy, která využívá energii buňky k transportu K+ do buňky a Na+ ven v poměru 2:3. V normálním stavu je membrána relativně propustná pro K+ (obsahuje permanentně otevřené K+ kanály), ale výrazně méně propustná pro Na+ a Cl−. Jelikož koncentrace K+ je v buňce vysoká a vně buňky nízká, difunduje K+ spontánně z buňky ven po koncentračním spádu, což vytváří záporný elektrický potenciál na vnitřní straně cytoplazmatické membrány vzhledem k prostředí vně buňky.[13]
Změny membránového potenciálu jsou umožněny pomocí specifických iontových kanálů v plazmatické membráně, které jsou schopné měnit propustnost membrány pro konkrétní ionty. Membránový potenciál většiny buněk je víceméně stabilní, jinak je to však u elektricky aktivních buněk (neuronů a svalových buněk).
Akční potenciál
editovatKlidový membránový potenciál neuronů je mezi -50 až -90 mV.[14] Při přijetí podnětu dojde k otevření iontových kanálů pro sodné ionty. Tyto ionty směřují po svém koncentračním i elektrickém gradientu směrem do buňky. Následkem toho se hodnoty membránového potenciálu dostávají blíže k nulové hodnotě (depolarizace membrány). Malé změny membránového potenciálu (a tedy změny napětí) mohou vést k otevření napěťově řízených iontových kanálů, a tím k dalšímu toku iontů přes membránu. Sníží-li se hodnota klidového potenciálu pod tzv. prahový potenciál, odpoví nervové vlákno vznikem akčního potenciálu. V další fázi (repolarizace) se propustnost membrány pro sodné ionty snižuje a potenciál se vrací ke klidovým hodnotám. Akční potenciál je tedy cyklus depolarizace a hyperpolarizace membrány a následného návratu ke klidovému potenciálu. Jeden akční potenciál trvá 1-2 ms. Akční potenciál se šíří jednostranně (díky inaktivaci Na+ kanálů po jejich aktivaci – tzv. refrakterní perioda) a bez úbytku.[12][13]
Přenos informace na synapsi
editovatAkční potenciál šířící se axonem do presynaptického zakončení axonu zde umožní otevření napěťově ovládaných vápníkových kanálů, což vede k toku vápenatých iontů do buňky. Zvýšená koncentrace vápenatých iontů v presynaptické části vede k interakcím proteinů přítomných v membráně presynaptického útvaru a v membráně synaptických vezikul, což následně vyústí ve splynutí presynaptické membrány se synaptickými vezikulami a vylití jejich obsahu do synaptické štěrbiny. Molekuly neuropřenašeče se ze synaptické štěrbiny vyvazují na receptory na postsynaptické membráně, čímž je informace přenesena do nové buňky. Zároveň je neuropřenašeč ze synaptické štěrbiny aktivně odstraňován zpětným vychytáváním do presynaptického neuronu, případně je degradován speciálními enzymy. Přenos informace na synapsi podléhá synaptickému zdržení (do 0,5 ms).[5]
Reference
editovat- ↑ a b c DEBANNE, Dominique; CAMPANAC, Emilie; BIALOWAS, Andrzej. Axon Physiology. Physiological Reviews. 2011-04-01, roč. 91, čís. 2, s. 555–602. Dostupné online [cit. 2019-07-26]. ISSN 0031-9333. DOI 10.1152/physrev.00048.2009.
- ↑ LUDĚK, Vajner; JIŘÍ, Uhlík; VÁCLAVA, Konrádová. Lékařská histologie I.: Cytologie a obecná histologie. [s.l.]: Charles University in Prague, Karolinum Press 113 s. Dostupné online. ISBN 9788024641072. Google-Books-ID: LIB1DwAAQBAJ.
- ↑ a b c d RADOMÍR, Čihák. Anatomie 3: Třetí, upravené a doplněné vydání. [s.l.]: Grada Publishing, a.s. 405 s. Dostupné online. ISBN 9788024756363. Google-Books-ID: A6mfCwAAQBAJ.
- ↑ August Newsletter: Actin Ring-based Periodic Membrane Skeleton in Neuronal Axons. www.cytoskeleton.com [online]. [cit. 2019-07-26]. Dostupné online.
- ↑ a b c d e MIROSLAV, Orel. Nervové buňky a jejich svět. [s.l.]: Grada Publishing, a.s. 217 s. Dostupné online. ISBN 9788024750705. Google-Books-ID: pZrGBwAAQBAJ.
- ↑ a b c d e SALZER, J. L.; ZALC, B. Myelination. Current Biology. 2016-10-24, roč. 26, čís. 20, s. R971–R975. Dostupné online [cit. 2019-07-26]. ISSN 0960-9822. DOI 10.1016/j.cub.2016.07.074.
- ↑ LÜLLMANN-RAUCH, Renate. Histologie. [s.l.]: Grada Publishing a.s. 578 s. Dostupné online. ISBN 9788024737294. Google-Books-ID: iMPSLaDsGyYC.
- ↑ JAROSLAV, Slípka; ZBYNĚK, Tonar. Základy histologie. [s.l.]: Karolinum Press 117 s. Dostupné online. ISBN 9788024628097. Google-Books-ID: VlKXBgAAQBAJ.
- ↑ a b c d e f MAHAR, Marcus; CAVALLI, Valeria. Intrinsic mechanisms of neuronal axon regeneration. Nature reviews. Neuroscience. 2018-6, roč. 19, čís. 6, s. 323–337. PMID 29666508 PMCID: PMC5987780. Dostupné online [cit. 2019-07-26]. ISSN 1471-003X. DOI 10.1038/s41583-018-0001-8. PMID 29666508.
- ↑ DALIBOR, Kučera. Moderní psychologie: Hlavní obory a témata současné psychologické vědy. [s.l.]: Grada Publishing a.s. 216 s. Dostupné online. ISBN 9788024746210. Google-Books-ID: je1mAQAAQBAJ.
- ↑ BLANQUIE, Oriane; BRADKE, Frank. Cytoskeleton dynamics in axon regeneration. Current Opinion in Neurobiology. 2018-08-01, roč. 51, čís. Cellular Neuroscience, s. 60–69. Dostupné online [cit. 2019-07-26]. ISSN 0959-4388. DOI 10.1016/j.conb.2018.02.024.
- ↑ a b ROSINA, Jozef; KOLÁŘOVÁ, Hana; STANEK, Jiří. Biofyzika pro studenty zdravotnických oborů. [s.l.]: Grada Publishing a.s. 218 s. Dostupné online. ISBN 9788024713830. Google-Books-ID: XZGg9qKgm0QC.
- ↑ a b DARNELL, James; BALTIMORE, David; MATSUDAIRA, Paul. The Action Potential and Conduction of Electric Impulses. Molecular Cell Biology. 4th edition. 2000. Dostupné online [cit. 2019-07-26]. (anglicky)
- ↑ JIŘÍ, Beneš; DANIEL, Jirák; FRANTIŠEK, Vítek. Základy lékařské fyziky. [s.l.]: Charles University in Prague, Karolinum Press 325 s. Dostupné online. ISBN 9788024626451. Google-Books-ID: PPOiCgAAQBAJ.
Externí odkazy
editovatObrázky, zvuky či videa k tématu axon na Wikimedia Commons